êô•{ ⦠΄≥éω
¹@ÉNÉçÉXÉgÉäÉfÉBÉEÉÄë°²Ά¹AÉOÉâÉĽzêΪ¨ô΄CêΪ²Χ½Lâη•E΄έ²ΧÉOÉ΄¹[Év²≈²Ζ¹B™yèκ²β™D²Χ²»²©²…âη•E²Χ²©²Ϋ²Ω²≈èZ²ώ²≈²Δ²Ρ¹Aëê²»²«²Τ²Τ²ύ²…êH²Ή²γ²ξ²Ρ™°ï®²Χ£Αä«²Χ£Ü²…™ϋ²Ν²Ρèoâη²ΒëùêB²Β²ή²Ζ¹Béü²Δ²≈ï≥ï÷²Τ²Τ²ύ²…îrüï²≥²ξ²Ρ²ή²Ϋ™yèκ²…΄A²ι²Τ²Δ²Λèz䬲π¨J²ηï‘²Β²Ρ²Δ²ή²Ζ¹B²±²Χâη•E²Άé΅äOêϋ²βèΝ™≈•ρ²ή²ΫäΘë΅²…ëΈ²Β²Ρîώèμ²…΄≠²Δ²Χ²≈¹AêΔäE£Ü²Χ²«²±²Χ™yèκ£Ü²…²ύ²Δ²ή²Ζ¹B²±²ΧÉNÉçÉXÉgÉäÉfÉBÉEÉÄë°²ΧçΉ΄έ²Σ¹Aèù¨ϊ²©²γëΧ™ύ²…™ϋ²Ν²Ϋ²η²ή²ΫêH﮲…ç§²Ε²Ν²ΡèΝâΜ䫲…™ϋ²Ν²Ϋ²η²Ζ²ι²Τïa΄C²π²®²±²Β²ή²Ζ¹B²±²ξ²πÉNÉçÉXÉgÉäÉfÉBÉEÉÄä¥êθè«²Τ²Δ²Δ²ή²Ζ¹B
¹@ëψï\™I²»ÉNÉçÉXÉgÉäÉfÉBÉEÉÄä¥êθ諲…²Ά¹AäOèùïîà ²©²γ™νïîëgêD²…΄έ²ΣêN™ϋ²Β²Ρ²®²±²ιÉKÉXâσαs²βîjèùï½²Τ¹AêH²Ήï®²…΄έ²Σ秙ϋ²Β²Ρ²®²±²ιÉEÉGÉ΄ÉVÉÖ΄έêH£Ü™≈諲ΤÉ{ÉcÉäÉkÉX£Ü™≈è«²Σ²†²η²ή²Ζ¹B²±²ξ²γ²ΧÉNÉçÉXÉgÉäÉfÉBÉEÉÄä¥êθ諲…²Ά¨¥àω΄έ²Σ™≈ëf²Τ¨Ρ²Έ²ξ²ι£`î£éΩ²πéYêΕ²Ζ²ι²±²Τ²ΣïKê{²≈²Ζ¹B²Τ²η²μ²·¹Aîjèùï½²Χ¨¥àω΄έ²ΣéYêΕ²Ζ²ιîjèùï½™≈ëf²Τ¹AÉ{ÉcÉäÉkÉX£Ü™≈è«²Χ¨¥àω²≈²†²ιÉ{ÉcÉäÉkÉX΄έ²ΣéYêΕ²Ζ²ιÉ{ÉcÉäÉkÉX™≈ëf²Ά΄…²Ώ²Ρ£véÄ™≈êΪ²Σ΄≠²≠¹Aîjèùï½™≈ëf²ΆëΧèd²P²΄²΅™•²η²Tng
(0.000005 mg)¹AÉ{ÉcÉäÉkÉX™≈ëf²Ά0.5 ng²≈™°ï®²πéE²Ζ²±²Τ²Σ²≈²Ϊ²ή²Ζ¹B²±²Χ™ώ²¬²Χ™≈ëf²Ά²Δ²Η²ξ²ύê_¨o¨n²…™≠²Δ²Ρâ^™°¨n²Χ•ÉαÉ²π²–²Ϊ²®²±²Β¹A¨Ρ΄zâ^™°²Σ²≈²Ϊ²»²≠²»²Ν²Ρ™°ï®²ΣéIJ…²ή²Ζ¹B²±²ξ²γ²Χ™≈ëf²…²φ²Ν²ΡÉ}ÉEÉX²Χ΄Ί™ς²Σ•ÉαÉ²Ζ²ιéά¨±²Χ½α²πê}²P²…éΠ²Β²ή²Β²Ϋ¹B²±²Χ™ώ²¬²Χê_¨o™≈ëf²Ά²Δ²Η²ξ²ύê_¨o²Χ΄Μï±²Σéü²§²ΧçΉ•E¹iê}²Χèξç΅²Ά΄Ί™ςçΉ•E¹j²…™`²Π²γ²ξ²ι²Χ²π•W²Α²ι™≠²Ϊ²π²ύ²Ν²Ρ²Δ²ή²Ζ¹B²±²ξ²πÉVÉiÉvÉX™`£B²ΧëOÉVÉiÉvÉXêΪ½}êß²Τ¨Ψ²Δ²ή²Ζ¹Bê}²Q²ΧÉXÉL¹[ÉIJΆ¹A²±²ξ²γ²Χ™≈ëf²Χçλ½p²≈²±²ξ²ή²≈²…âπ²Ν²Ρ²Δ²ι²±²Τ²Χ²ή²Τ²Ώ²≈²Ζ¹Bîjèùï½™≈²®²φ²―É{ÉcÉäÉkÉX™≈ëf²ΆÉVÉiÉvÉX²Χê_¨oèI••ëΛ²…¨΄ç΅²Β¹Aê_¨oèI••ïîïΣ²…éφ²ηçû²ή²ξ²ή²Ζ¹B²¬²Δ²≈¹A™≈ëf²Χ¨yçΫ²ΤèdçΫ²Χâπ½Θ²Σ²®²±²η¹B¨yçΫïîïΣ²ΣÉVÉiÉvÉX觕E²ΤÉVÉiÉvÉXëO•¨²Τ²Χ½Zç΅²…ä÷¨W²Ζ²ιSNARE£`î£éΩ²π¨ά£ηïΣâπ²Β²ή²Ζ¹BSNARE£`î£éΩ²ΣïΣâπ²≥²ξ²Ϋê_¨oèI••²≈²ΆÉVÉiÉvÉX觕E²ΤÉVÉiÉvÉXëO•¨²Σ½Zç΅²≈²Ϊ²»²≠²»²ι²Χ²≈¹AÉVÉiÉvÉX觕E²…£~²Π²γ²ξ²Ρ²Δ²ιê_¨o™`£Bï®éΩ²ΣÉVÉiÉvÉX䑨³²…ïζèo²≥²ξ²»²≠²»²ηê_¨o•ÉαÉ²Σ²®²±²η²ή²Ζ¹B
¹@²Μ²ξ²≈²Ά¹Aîjèùï½²ΤÉ{ÉcÉäÉkÉX£Ü™≈諲Ά™·²Ε²»²Χ²©²Τ¨Ψ²Λ²Τ¹AëS²≠è«èσ²Άê≥îΫëΈ²»²Χ²≈²Ζ¹Bîjèùï½²≈²ΆëSêg²Χç€äi΄Ί²Σçd²≠éϊèk²Β²Ρ•ÉαÉ²Ζ²ι²Χ²…ëΈ²Β²Ρ¹AÉ{ÉcÉäÉkÉX£Ü™≈諲≈²Άç€äi΄Ί²Σä…²ώ²≈éϊèk²Β²»²≠²»²ι²Χ²≈²Ζ¹B²±²Χàα²Δ²Ά¹AÉ{ÉcÉäÉkÉX™≈ëf²Σéε²…••èΫ²Χê_¨o΄Ί™`£B²πëjäQ²Ζ²ι²Χ²…ëΈ²Β²Ρ¹Aîjèùï½™≈ëf²Σ£Üêï²Χâ^™°ê_¨oçΉ•E²…ëΈ²Ζ²ιÉVÉiÉvÉX²πéε²…ëjäQ²Ζ²ι²Ϋ²Ώ²Ψ²Τçl²Π²γ²ξ²Ρ²Δ²ή²Ζ¹B
¹@²Μ²ξ²≈²Ά¹A²±²ξ²γ²Χ™≈ëf²Ά••èΫ²Τ£Üêï²Τ²π²«²Χ²φ²Λ²…²Β²Ρé·ï ²Β²Ρ²Δ²ι²Χ²≈²Β²ε²Λ²©¹H²±²Χ™ö²Π²Ά²ή²Ψ•Ψ²γ²©²…²≥²ξ²Ρ²Δ²ή²Ι²ώ¹B²ή²Ϋ¹Aîjèùï½™≈ëf²Σ²«²Χ²φ²Λ²…²Β²Ρ£Üêïê_¨o¨n²ή²≈â^²Έ²ξ²ι²Χ²©²…²¬²Δ²Ρ²ύ™ö²Σèo²Ρ²Δ²ή²Ι²ώ¹B²±²ξ²γ²Χ΄^•β²…™ö²πèo²Ζ²±²Τ²Σé³²Χ•≤²Χà²≈²Ζ¹B
|
|
ê}²P¹Dîjèùï½™≈ëf²…²φ²ιê_¨o΄Ί΄Μï±™`£BëjäQ¹BÉ}ÉEÉX²ΧâΓäu•¨²πâΓäuê_¨o²Τ²Τ²ύ²…éφ²η²Ψ²Β²Ρ¹AâΓäuê_¨o²…ëΈ²Ζ²ι™d΄Céh¨É²…²φ²Ν²Ρ½Uî≠²≥²ξ²ιâΓäu•¨²Χéϊèk£Θ½Ά²π¨oéû™I²…΄L‰^²Β²Ϋ¹Bîjèùï½™≈ëf²πçλ½p²≥²Ι²ι²Τ¹A™·²Ε΄≠²≥²Χéh¨É²≈½Uî≠²≥²ξ²ιéϊèk£Θ½Ά²Σ²Β²Ψ²Δ²…éψ²≠²»²Ν²Ρ²Δ²≠²Χ²ΣîF²Ώ²γ²ξ²ι¹BâΚ²≈¹Aê_¨o΄Ί™`£B²…ëΈ²Ζ²ιîjèùï½™≈ëf²ΤÉ{ÉcÉäÉkÉX™≈ëf²ΧîZ™x¹]çλ½p£Φêϋ²Χîδär¹BÉ{ÉcÉäÉkÉX™≈ëf²Χïϊ²Σê_¨o΄Ί™`£B²…ëΈ²Β²Ρ²Ά΄≠²Δçλ½p²πéù²Ν²Ρ²Δ²ι²±²Τ²Σâπ²ι¹B |
|
|
ê}²Q¹Dîjèùï½�≈ëf²ΤÉ{ÉcÉäÉkÉX™≈ëf²Χçλ½p΄@ç\²ΧÉX�L¹[ÉĹBê_¨o™`£Bï®éΩ²Σ™ϋ²Ν²Ρ²Δ²ιÉVÉiÉvÉX觕E²Χsynaptobrevin (VAMP)²ΤÉVÉiÉvÉXëO•¨²…ëΕçί²Ζ²ιSNAP-25²®²φ²―syntaxin²ΣNSF²βÉΩÉάÉΝSNAP²πâν²Β²ΡïΓç΅ëΧ²π¨`ê§²Ζ²ι²Ζ²ι²±²Τ²≈¹AÉVÉiÉvÉX觕E²ΣÉVÉiÉvÉXëO•¨²…¨≈£η²≥²ξ²ι¹B¨≈£η²≥²ξ²Ϋ觕E²ΆÉJÉ΄ÉVÉEÉÄîZ™xàΥëΕêΪ²…ÉVÉiÉvÉXëO•¨²Τ½Zç΅²Β¹A觕E™ύ²Χ™`£Bï®éΩ²ΣÉVÉiÉvÉX䑨³²…ïζèo²≥²ξ²ι²±²Τ²≈ê_¨o™`£B²Σ²»²≥²ξ²ι¹Bîjèùï½™≈ëf²ΤÉ{ÉcÉäÉkÉX™≈ëf²Χ¨yçΫ²Ά²±²ξ²γ²ΧSNARE£`î£éΩ²π¨ά£ηïΣâπ²Ζ²ι²±²Τ²≈ÉVÉiÉvÉX觕E²ΤÉVÉiÉvÉXëO•¨²Τ²Χ½Zç΅²πëjäQ²Β¹A™`£Bï®éΩ²Σïζèo²≥²ξ΄Μï±²Σ™`²μ²ι²±²Τ²πé’£f²Ζ²ι¹B |
à…ê®êλ ½T™ώ èy΄≥éω
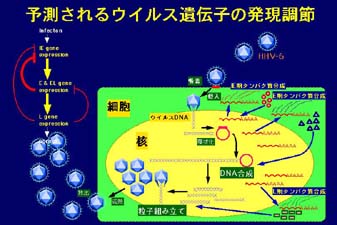
¹@ÉqÉgÉwÉ΄ÉyÉXÉEÉCÉ΄ÉX²U(HHV-6)²Ά²QçΈ²ή²≈²…90%à»èψ²Χ½céô²Σä¥êθ²Β¹A²Μ²ΧÉEÉCÉ΄ÉX²πéùë±ä¥êθ²ύ²Β²≠²Άêωïöä¥êθ²Χ¨`²≈àξêΕéù²Ωë±²·¹Aéû¹XçΡäàêΪâΜ²≥²ξ²Ρ¹A²®²φ²Μ30%²Χê§êl²Χswab²©²γgenome
DNA²Σ¨üèo²≥²ξ²ι¹B²ή²Ϋ¹AHHV-6²Άïξéqä¥êθ²…²φ²η™`îά²≥²ξ²ι¹B
¹@HHV-6²Χä¥êθ¨ψ¹Aäj™ύ²…genome DNA²πë}™ϋ²Β¹Agenome DNA²Άä¬èσâΜ²Χ¨ψ¹Alytic²»¨o‰H²†²ι²Δ²Άêωïöä¥êθ¨o‰H²…²Ά²Δ²ι²Τçl²Π²γ²ξ²Ρ²Δ²ι¹B²Β²©²Β¹A¨Μçί²Χ²Τ²±²κlytic²»¨o‰H²Τêωïö¨o‰H²Τ²ΧêΊ²ηë÷²Π²Χ΄@ç\²βêωïöÉEÉCÉ΄ÉX²©²γ²ΧçΡäàêΪâΜ²Χ΄@ç\²…ä÷²Β²Ρ²ΆëS²≠£m²γ²ξ²Ρ²Δ²»²Δ¹B²®²Μ²γ²≠èhéεçΉ•E²…™ϋ²ιÉVÉOÉiÉ΄²ΧÉlÉbÉgÉè¹[ÉN²π½‰½p²Β¹A²Μ²ξ²γ²ΧêΊ²ηë÷²Π²πçs²Ν²Ρ²Δ²ι²ύ²Χ²Τ½\ëΣ²≥²ξ²Ρ²Δ²ι¹B
¹@HHV-6²Χä¥êθ²Ζ²ιçΉ•E²Ά²sçΉ•E¹Amonocyte/macrophage¹ANKçΉ•E²≈²†²ι²±²Τ²©²γ¹A•Τâu¨n²…™ϋ²ιÉVÉOÉiÉ΄™`£B¨o‰H²πHHV-6²Σ½‰½p²Ζ²ι²±²Τ²Σ½\ëΣ²≥²ξ¹AHHV-6²Σ•Τâu¨n²Χêß¨δ²…²ύä÷½^²Ζ²ιâ¬î\êΪ²Σ²†²ι¹B²≥²γ²…¹AHHV-6ä¥êθ²…²φ²ιàξïî²ΧÉTÉCÉgÉJÉCÉ™²Χ°²Άäυ²…ïώçê²Β¹AëΦ²ΧÉOÉ΄¹[Év²©²γ²ύïώçê²≥²ξ²Ρ²Δ²ι²Σ¹AÉTÉCÉgÉJÉCÉ™ÉlÉbÉgÉè¹[ÉNëSëΧ²Χ½ùâπ²…²Ά΄y²ώ²≈²Δ²»²Δ¹B
¹@HHV-6²…²Άvariant A²ΤB²ΣëΕçί²Β¹Avariant
B²Ά™Υî≠êΪî≠ê]²Χ¨¥àωÉEÉCÉ΄ÉX²≈²†²ι²Τïώçê²≥²ξ²Ϋ²Σ¹Avariant A²Χïa¨¥êΪ²Άïώçê²≥²ξ²Ρ²®²γ²Η¹A²ή²Ϋ¹Avariant A²ΧU1102äî²ΧëSâ•äνîz½ώ²ΆGompels²γ²…²φ²η²Ζ²≈²…ïώçê²≥²ξ²Ϋ²Σ¹Avariant
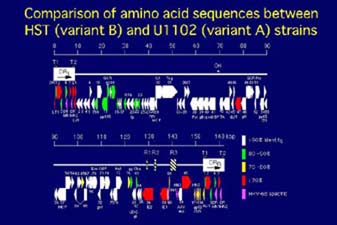
B²ΧHSTäî²ΧëSâ•äνîz½ώ²Άâδ¹X²Σ¨à£η²Β²Ϋ¹iê}²P¹j¹B½Φvariant 䑲≈²Χâ•äνîz½ώ²πîδär²Β²Ϋ²Τ²±²κ¹Aëε²Ϊ²≠àΌ²»²ιïîà ²Σ½Φ£[²Χdirect repeat²Χïîà ²Τunique½Χàφ²Χ²R²¬²Χ¨J²ηï‘²Β½Χàφ²≈²†²Ν²Ϋ¹B½\ëΣ²≥²ξ²ιopen
reading frame (ORF)²Άdirect repeat™ύ²…10¹Aunique½Χàφ²…88ëΕçί²Β²Ϋ¹BDirect repeat™ύ²ΧORF²Άvariant䑲≈ëε²Ϊ²≠àΌ²»²η¹Avariant™ΝàΌ™I²»OFR²ύëΕçί²Ζ²ι¹BUnique½Χàφ²ΧORF²ΆëSëΧ™I²…²Ά²®²φ²Μ95%²Χëä™·êΪ²πéΠ²Β²Ϋ²Σ¹A²W²¬²ΧORF²Ά80-90%¹A²W²¬²ΧORF²Ά70-80%¹A²S²¬²ΧORF²Σ70%à»âΚ²Χëä™·êΪ²≈²†²Ν²Ϋ¹B²±²ξ²γ²Χàβ™`éq²Χëäàα²Σïa¨¥êΪ²βä¥êθêΪ²Χàα²Δ²…ä÷‰A²Β²Ρ²Δ²ι²Χ²©²Χ²ύ²Β²ξ²»²Δ¹B²±²Χ²φ²Λ²»àβ™`éqâπêΆ²Χ£Ü¹Aä¥êθçΉ•E²ΧÉVÉOÉiÉ΄™`£B²Χä÷½^²Ζ²ι²Τ½\ëΣ²≥²ξ²ιàβ™`éq²π²R²¬
(U12, 51, 83) ¨©²Δ²Ψ²Β²Ϋ¹BU12, 51²ΆG-protein coupled receptor (GCR) homolog²≈²†²η¹AU83²Άchemokine
homolog²≈²†²Ν²Ϋ¹B
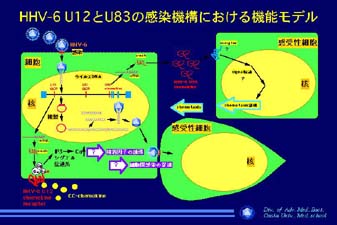
¹@U12²ΤU83²Χ΄@î\âπêΆ²πçs²Ν²Ϋ¨΄â ¹AU12²ΆCC-chemokine receptor²≈²†²ι²±²Τ¹AU83²Άviral chemokine²≈²†²η¹Amonocyte²…ëΈ²Ζ²ιchemoattractantî\²π½L²Β²Ρ²Δ²ι²±²Τ²π•Ψ²γ²©²…²Β²Ϋ¹iê}2¹j¹BDNA
chip²π½p²Δ²Ρàβ™`éq²Χî≠¨ΜÉpÉ^¹[É™²ύ•Ψ²γ²©²…²Β¹AÉJÉXÉP¹[Éh²Χ£≤êΏ΄@ç\²ΧëΕçί²ΣéΠ祲≥²ξ²Ϋ¹iê}²R¹j¹B£Ü‰açRëΧ²ΧîFé·²Ζ²ι™€£`ΧîFé·²Ζ²ιÉAÉ~Émé_²πHHV-6A²ΤB²ΧÉLɹÉâ£`©²γ¨à£η²Β²Ϋ¹B¨Μçί¹CHHV-6²Χàβ™`éq£≤êΏ΄@ç\²ΤÉTÉCÉgÉJÉC²Χî≠¨Μ£≤êΏ΄@ç\²Χâπ•Ψ²…ëS½Ά²πèψ²Α²Ρéφ²ηëg²ώ²≈²Δ²ι¹B
|
|
ê}²P¹D |
|
|
ê}²Q¹D |
|
|
ê}²R¹D |
¨Υγ≥ ΄€ èy΄≥éω
¹@é³²Ϋ²Ω²Χ¨Λ΄ÜéΚ²≈²ΆëΫ²≠²Χïa¨¥êΪëε£Α΄έ²Χ²Λ²Ω™Ν²…²Qéμ½ό²Χëε£Α΄έ²…²¬²Δ²Ρ¨Λ΄Ü²πçs²Ν²Ρ²Δ²ή²Ζ¹B à²Ά£Αä«èo¨¨êΪëε£Α΄έ¹iEHEC¹j²≈éε²…èo¨¨êΪëε£Αâä²βîA™≈è«è«¨σ¨Q²»²«²πàχ²Ϊ΄N²±²Β²ή²Ζ¹B²P²X²X²UîNçδés™ô™ζ•{äe£n²≈ëε²Ϊ²»îμäQ²πèo²Β²ΫO157:H7²Ά²±²ξ²Χëψï\™I²»¨¨ê¥¨^²≈²Ζ¹BEHEC
O157²Χïa¨¥êΪ²…²Άéuâξ™≈ëféYêΕî\²Σèd½v²»•πä³²π£S²Ν²Ρ²Δ²ή²Ζ²Σ¹AÉqÉg£Αä«èψîγçΉ•E²÷²Χït£Öî\²ύèhéε¹AïW™IëgêDîFé·¹A£Αä«™ύ²≈²ΧëùêB²…ïKê{²≈²†²ι²±²Τ²Ά•Ψ²γ²©²…²»²Ν²Ρ²Δ²ή²Ζ¹BEHEC²ΆèhéεçΉ•E²…ït£Ö²Ζ²ι²±²Τ²…²φ²ηèhéεçΉ•E²…½l¹X²»èαäQ²πàχ²Ϊ΄N²±²Β²ή²Ζ¹B
¹@²ή²Ϋ²ύ²Λà²Χ¨Λ΄ÜëΈèέ²Τ²Β²Ρ²Δ²ι£Αä«ïa¨¥êΪëε£Α΄έ¹iEPEC¹j²Ά²β²Ά²ηÉqÉg²…âΚ½ü²πàχ²Ϊ΄N²±²Β²ή²Ζ²Σ¹A™≈ëféYêΕî\²Ά•≥²≠çΉ•Eït£Ö²…²φ²ιçΉ•EèαäQ²Σéε²»ïa¨¥êΪ²≈²†²ι²Τçl²Π²γ²ξ²Ρ²Δ²ή²Ζ¹BEHEC²ΧçΉ•Eït£Ö½lé°²Χàξïî²ΆEPEC²Χ½lé°²Τ΄Λ£ ²≈²Ζ²Σ¹AçΉ•Eït£Ö²Χèâäζ£iäK²Σëε²Ϊ²≠àΌ²»²Ν²Ρ²Δ²ι²±²Τ²Σ•Ψ²γ²©²…²»²Ν²Ρ²Δ²ή²Ζ¹B
¹@é³²Ϋ²Ω²Ά²±²ξ²γ²ΧçΉ•Eït£Ö²…ä÷²μ²ιàωéq²ΧéάëΧ²ΤèhéεçΉ•E²Τ²Χëä¨ίçλ½p²πïΣéqêΕï®äw™I¹AçΉ•EêΕï®äw™Iéη•@²π½p²Δ²Ρâπ•Ψ²Β²φ²Λ²Τ²Β²Ρ²Δ²ή²Ζ¹B²ή²Ϋ¹A΄ΏîN¨à£η²≥²ξ²ΫEHEC
O157 Sakaiäî²ΧëSÉQÉmÉÄâ•äνîz½ώèνïώ²…äν²Ο²Ϊïa¨¥êΪä÷‰Aàβ™`éq¨Q²Χ•‘½Ö™I¨üçθ²®²φ²―î≠¨ΜÉlÉbÉgÉè¹[ÉN²ΧâπêΆ²πäJén²Β²Ρ²®²η²ή²Ζ¹B
|
|
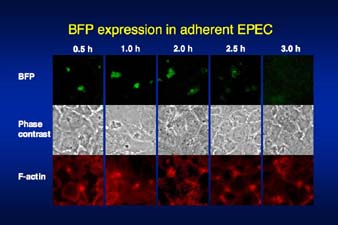
ê}²P ¹D£Αä«ïa¨¥êΪëε£Α΄έEPEC²ΧèψîγçΉ•Eït£Ö¨ψ²Χîςè§èW½é¨`꧹i£Ü£i¹jâΏ£ω²…²®²·²ιë©èσêϋ•―¹iBFP,èψ£i¹jî≠¨Μ½}êß²ΤÉAÉNÉ`É™΄ΟèW¹iâΚ£i¹j½U™±²Χéûä‘™IïœâΜ |
|
|
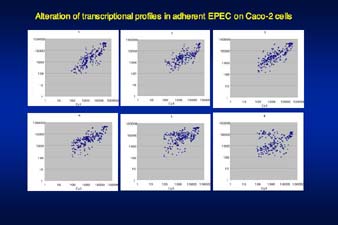
ê}²Q¹DèψîγçΉ•E²…ït£Ö²Β²ΫEPEC²…²®²·²ιàβ™`éq™]é ²Χ¨oéû™IïœâΜ²πDNA microarray²π½p²Δ²ΫâπêΆ |