II型膜タンパク質CKAP4の機能
パルミチン酸化CKAP4によるVDAC2を介したミトコンドリア機能の制御
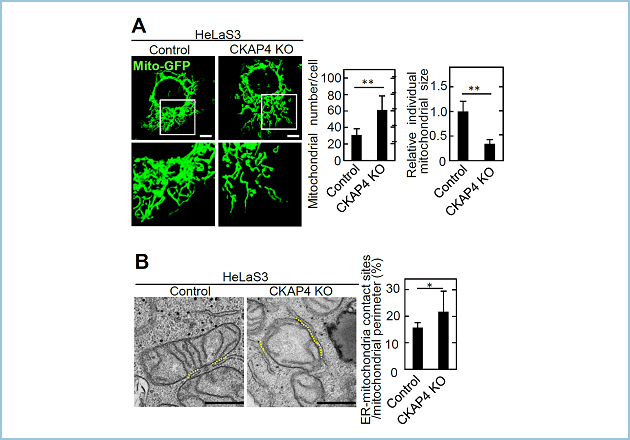
CKAP4 (cytoskeleton-associated protein 4)は主として小胞体に局在し、N端側を細胞質側に、C端側を小胞体内腔側に向けるII型膜タンパク質であり、小胞体の構造維持に関与することが報告されています。今回私共は、CRISPR/Cas システムを用いて作製したCKAP4ノックアウト(KO)細胞において、これまでに報告されている小胞体の構造異常に加えて、ミトコンドリアの断片化および小胞体ーミトコンドリア接触点の増加が生じることを見出しました(図1)。
図1.CKAP4 KO細胞におけるミトコンドリアの構造および小胞体ーミトコンドリア接触点の異常
(A) ControlおよびCKAP4 KO細胞にMito-GFPを発現させ、ミトコンドリアの構造を観察した。
(B) ControlおよびCKAP4 KO細胞の小胞体ーミトコンドリア接触点(黄色点線)を電子顕微鏡により観察した。
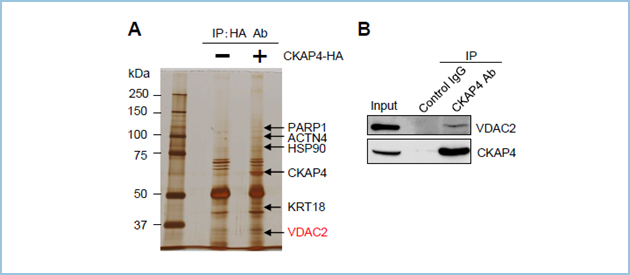
しかし、主に小胞体に局在するタンパク質であるCKAP4がミトコンドリアの構造を制御するメカニズムは不明でした。そこでCKAP4の新規結合タンパク質を探索し、ミトコンドリア外膜に存在するチャネルタンパク質VDAC2を見出しました(図2)。
図2.CKAP4新規結合タンパク質の探索と同定
(A) HA抗体を用いた免疫沈降によるCKAP4新規結合タンパク質候補の同定。
(B) CKAP4はVDAC2と特異的に結合する。
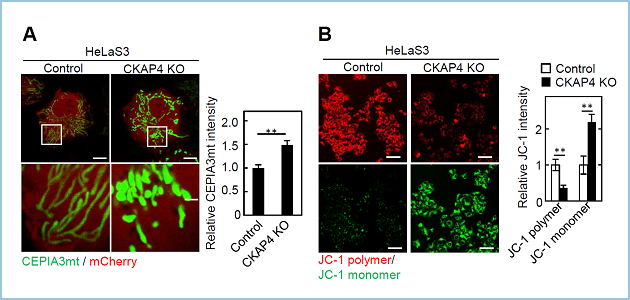
VDAC2は小胞体ーミトコンドリア接触点において、イノシトール三リン酸受容体(IP3R)やGRP75と相互作用し、小胞体からミトコンドリアへのCa2+の受け渡しに関わることが報告されています。生化学的解析および免疫電子顕微鏡観察によりCKAP4の一部はVDAC2と同様にMAMに局在することを明らかにしました。CKAP4とVDAC2の結合には、CKAP4の細胞質領域に存在する100番目のシステイン残基 (Cys100)のパルミチン酸化が必要でした。CKAP4 KO細胞ではVDAC2とIP3Rの相互作用が増強することにより、ミトコンドリア内へのカルシウムの流入とそれに伴うミトコンドリアの膜電位の低下が観察されました(図3)。
図3.CKAP4 KO細胞におけるミトコンドリア内カルシウム濃度およびミトコンドリア膜電位の異常
(A) ControlおよびCKAP4 KO細胞にミトコンドリア特異的カルシウムインディケーターCEPIA3mtを発現させ、ミトコンドリア内のカルシウム濃度を測定した。
(B) JC-1によるミトコンドリアの膜電位の測定。
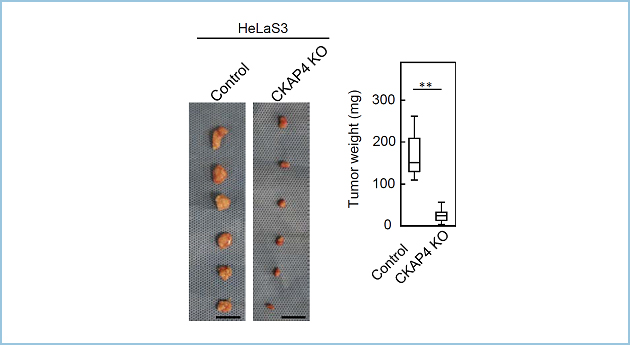
さらにCKAP4 KO細胞はミトコンドリアの呼吸と低グルコース条件下での細胞増殖が低下し、ヌードマウスを用いたゼノグラフトモデルにおいて皮下腫瘍の形成能が低下しました(図4)。
図4.ゼノグラフトモデルにおけるCKAP4 KO細胞の腫瘍形成能の低下
ヌードマウスの皮下に細胞を移植するゼノグラフトモデルを用いて、ControlおよびCKAP4 KO細胞の皮下腫瘍の形成能を評価した。
これらのCKAP4 KO細胞におけるミトコンドリア機能の異常は野生型CKAP4の発現によりレスキューされましたが、パルミチン酸化欠損型CKAP4変異体の発現によってはレスキューされませんでした。以上の結果から、パルミチン酸化CKAP4はVDAC2と結合し、ミトコンドリアへのカルシウム流入を制御することによりがん細胞の増殖に関与することが示唆された。今後はCKAP4によるミトコンドリアの機能制御の生理的意義について個体レベルで明らかにしていく予定である。
細胞膜CKAP4による細胞運動制御
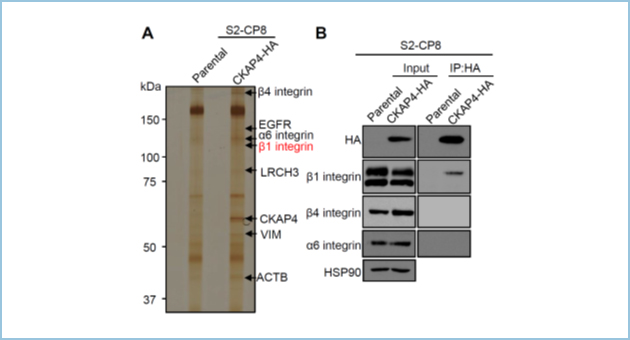
私達の研究室では、Wnt/β-cateninシグナルの構成因子であるDickkopf1(DKK1)の新規受容体としてCytoskeleton-associated protein 4 (CKAP4)を同定しました(Wntシグナルと癌を参照)。DKK1-CKAP4シグナルはphosphatidylinositol-3 kinase(PI3K)-AKT経路を活性化して、がん細胞の増殖を促進します。同時に、CKAP4とDKK1は細胞運動も促進しますが、その機序は不明でした。そこで私共は、CKAP4による細胞運動制御機構を明らかにすることを試みました。様々ながん細胞株を用いて解析したところ、細胞膜上CKAP4はリガンドであるDKK1と独立して、細胞運動と細胞運動時の接着斑ターンオーバーを促進することが判明しました。また、CKAP4はβ1インテグリンと結合しました(図1)。
図1.CKAP4新規結合タンパク質の探索と同定
(A) 膵がん細胞株S2-CP8細胞においてCKAP4新規結合タンパク質候補としてインテグリンファミリーの同定。
(B) CKAP4はβ1インテグリンと特異的に結合する。
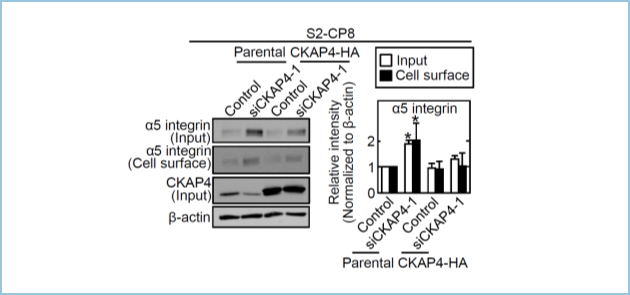
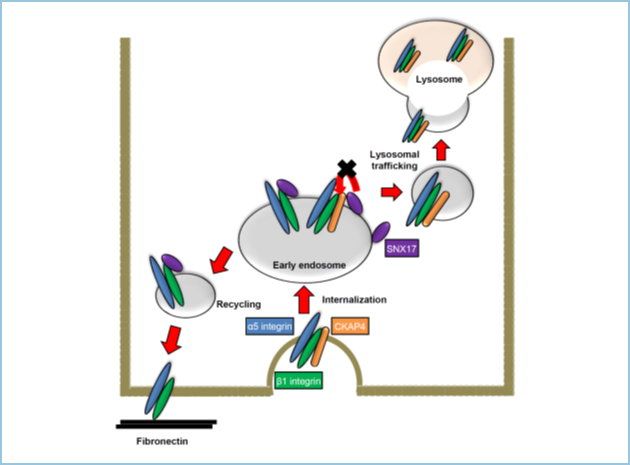
CKAP4ノックダウンは、β1インテグリンのパートナーであるα5インテグリンの発現量をタンパク質レベルで増加させました(図2)。細胞膜CKAP4は、β1インテグリンと結合し、早期エンドゾーム上でSorting nexin (SNX17)のβ1インテグリンへの結合と競合することにより、α5β1インテグリンのリサイクリングを抑制し、その結果、α5β1インテグリンのリソソームへの輸送と分解を促進しました。すなわち、細胞膜CKAP4はDKK1シグナルとは独立して、細胞膜上のα5β1インテグリンの発現を適切に抑制することにより、細胞運動を促進することが示唆されました(図3)。
図2.CKAP4によるα5インテグリンの制御
CKAP4ノックダウンにより、α5インテグリンのタンパク質レベルが増加した。
図3.細胞膜上CKAP4によるα5β1インテグリンのリサイクリングの制御
α5β1インテグリンと複合体を形成した細胞膜上CKAP4はエンドサイトーシスされ後、早期エンドソーム上でSNX17とβ1インテグリンとの結合に競合することで、α5β1インテグリンのリサイクリングを抑制する。その結果、α5β1インテグリンはリソソームに輸送され、分解される。