Research Projects
We aim to figure out that roles of signal transduction pathway in cellular growth, differentiation, motility, and polarity to understand their disruption-mediated diseases and develop novel diagnostic and therapeutic tools. Although we particularly focus on Wnt signaling pathway, we are also analyzing other pathways. We recognize the signal transduction itself as a biological system that links genotypes with phenotypes, and we will create new paradigm of life science research and the understanding of diseases. The following studies are under the way.
INDEX
1. Wnt signaling and cancer
Arl4c regulates pancreatic cancer invasion through invasive pseudopods in an invadopodia-independent pathwayNew
Pancreatic cancer is extremely aggressive and has a high mortality rate due to metastasis. Whereas KRas is mutated in most pancreatic cancer patients, controlling KRas or its downstream effectors has not yet been succeeded clinically.
Arl4c is a small G protein of which expression is induced by KRas and Wnt signaling and plays an important role in epithelial morphogenesis.
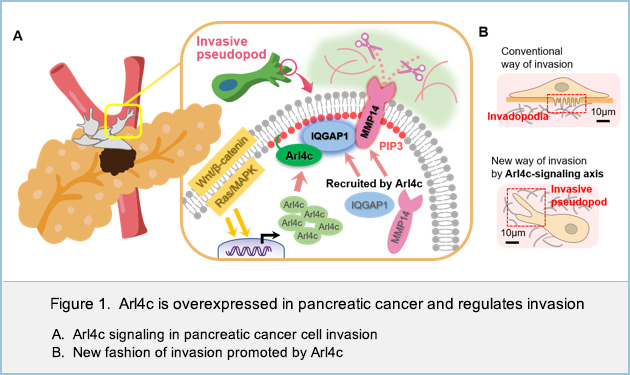
In this study, we found that Arl4c is frequently overexpressed in pancreatic cancer where both KRas and Wnt pathways are activated and that its localization to the tips of invasive pseudopods is required for cancer cell invasion.
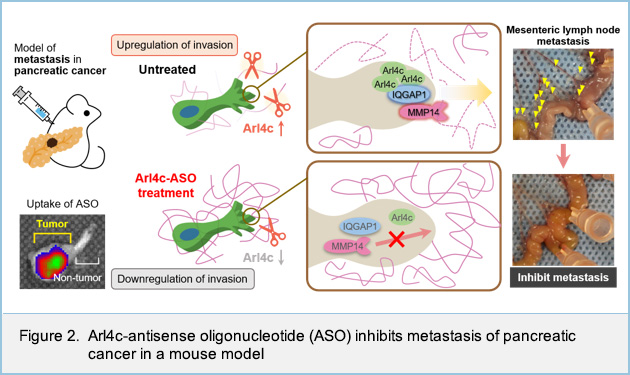
Furthermore, we showed that Arl4c interacts with IQGAP1, a scaffold protein for signaling pathway. Arl4c recruited IQGAP1 and its downstream effector, MMP14, to invasive pseudopods especially in the phosphatidylinositol (3,4,5)-trisphosphate (PIP3) enriched domain. Arl4c, IQGAP1 and MMP14 were specifically localized in the active site of invasion and induced degradation of extracellular matrix (Figure 1A). Although Arl4c was not involved in the formation of invadopodia, a well-known invasive structure at the ventral sites of cells, our new model with invasive pseudopods at the front end of cells shows how Arl4c promotes invasion in pancreatic cancer (Figure 1B).
In addition, using the mouse model, we showed that the inhibition of Arl4c expression by antisense oligonucleotide directly suppresses the invasive ability of pancreatic cancer cells (Figure 2).
In conclusion, this study clarified that invasion of pancreatic cancer cells is promoted by Arl4c, of which expression is induced by KRas and Wnt signaling, and that the association of Arl4c with IQGAP1 and MMP14 at the tips of invasive pseudopods in a PIP3-dependent manner is essential for the invasive ability. Arl4c might represent an appropriate target for pancreatic cancer therapy, as targeting KRas effector signaling pathway.
The positive feedback loop in the DKK1-CKAP4-FOXM1 axis is associated with poor prognosis in human pancreatic esophageal cancerNew
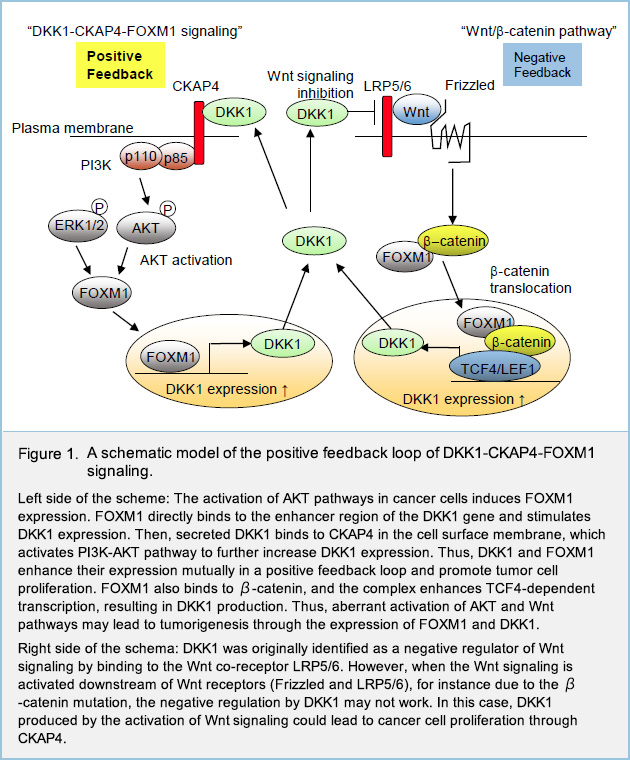
Recently, we have identified CKAP4 as a novel receptor for the secreted glycoprotein Dickkopf1 (DKK1), a Wnt antagonist, and also reported that DKK1-CKAP4 signaling promotes cancer cell growth via activation of the phosphoinositide 3-kinase (PI3K)-AKT pathway. Active DKK1-CKAP4 signaling promotes cancer cell proliferation and is associated with poor prognosis in various cancers including pancreatic ductal adenocarcinoma (PDAC), lung cancer, and esophageal squamous cell carcinoma (ESCC), but it remains unclear which genes and signaling pathways are involved downstream of DKK1-CKAP4 signaling. Moreover, although DKK1 expression is regulated downstream of the Wnt/β-catenin pathway, sometimes it is overexpressed in tumors in which Wnt/β-catenin pathway was not activated, but the underlying mechanism is yet unknown.
To find the uncharacterized signaling pathway regulated downstream of DKK1-CKAP4 signaling, we generated human PDAC cell line S2-CP8 in which endogenous DKK1 was knocked out, and then RNA sequencing analyses were performed using these cells and control S2-CP8 cells, and we focused on transcription factor FOXM1, which is known as a master regulator of cell cycle, and also known as an oncogene overexpressed in many cancerous diseases. In practice, expression levels of FOXM1 tended to be high in several PDAC and ESCC cell lines which expressed both DKK1 and plasma membrane CKAP4 (i.e. DKK1-CKAP4 signaling was activated). Knockdown of DKK1 or CKAP4 in these cells reduced FOXM1 expression. In addition, in vitro assay using AKT inhibitor revealed that FOXM1 expression was regulated downstream of DKK1-CKAP4 signaling via AKT activation. Besides, we found that the expression levels of DKK1 was fluctuated in parallel with FOXM1 expression levels. These results suggest that DKK1-CKAP4 signaling is involved in FOXM1 expression via AKT activation, and that FOXM1 expression is required for DKK1 expression.
Actually, Knockdown of FOXM1 in human PDAC and ESCC cell lines decreased the expression of DKK1, while overexpression of FOXM1 in human PDAC cell Capan-1, which expresses neither DKK1 nor FOXM1 endogenously, increased the expression of DKK1, which suggested that DKK1 is a target gene of transcription factor FOXM1
.
Then, we explored the 5' upstream region of the DKK1 gene in the genome and found consensus sequences of FOXM1 in about 2000 base pairs upstream from the transcription start site of the DKK1 gene (FOXM1 binding site). Chromatin immunoprecipitation assay using S2-CP8 cells confirmed the FOXM1 binding to the FOXM1 binding site. Then, S2-CP8 cells in which FOXM1 binding site is deleted from the genome was generated using Crispr-Cas9 system (S2-CP8/FOXM1 BS deletion (ΔFOXM1 BS) cells). As expected, DKK1 expression was decreased by depleting the FOXM1 BS. Moreover, both the cell proliferative activity in vitro and tumor formation in subcutaneous xenograft tumor models of S2-CP8/ΔFOXM1 BS cells were reduced than those of control cells. These results indicate that FOXM1 directly binds to the 5' upstream region of the DKK1 gene and stimulates DKK1 expression, which forms a positive feedback loop between DKK1 and FOXM1 to promote cancer cell proliferation (Fig. 1).
We then analyzed the correlation between DKK1 and FOXM1 expression in clinical cases of human PDAC and ESCC by Immunohistochemistry (IHC) study using surgically resected specimens. IHC staining for DKK1 and FOXM1 in PDAC tissues (38 cases) and ESCC tissues (82 cases), who underwent surgical resection at Osaka University hospital, was performed. In these cases, 27/38 (71.1%) PDAC cases and 40/82 (48.8%) ESCC cases were positive for both DKK1 and FOXM1 in serial sections. The correlation between the ratios of tumor lesions with positive DKK1 and FOXM1 staining was statistically significant (Fig. 2). The relapse-free survival rate tended to be lower in PDAC cases with the expression of both DKK1 and FOXM1 compared to other cases. In addition, the TCGA dataset (174 cases) indicated a significant correlation between the expression of FOXM1 and DKK1 mRNAs in PDAC cases, and when cases were separated into DKK1 and FOXM1 positive expression group and others, overall survival was significantly reduced in the DKK1 and FOXM1 positive expression group (Fig. 2). In ESCC cases, both the relapse-free survival rate and overall survival rate were significantly poor in DKK1 and FOXM1 double positive cases (Fig. 2). Taken together, these results indicate that the simultaneous expression of DKK1 and FOXM1 is associated with poor prognosis in PDAC and ESCC.

In conclusion, we elucidated that the transcription factor FOXM1 is expressed in human PDAC and ESCC cells downstream of DKK1-CKAP4 signaling through activated AKT, and that FOXM1 binds to the enhancer region of DKK1 and induce its expression, which forms a positive feedback loop to promote mutual expression and results in poor clinical prognosis.
Arl4c is associated with initiation and progression of lung adenocarcinoma and represents a molecular target for lung cancer therapy
ADP-ribosylation factor-like 4c (Arl4C), a member of ARL family proteins, is a target protein that is expressed by the activation of Wnt/β-catenin and EGF/RAS/mitogen-activated protein kinase (MAPK) signaling. We have previously reported Arl4c expression in human lung adenocarcinoma cell lines was important for the motility, invasion, and proliferation of cancer cells. In addition, we have developed an antisense oligonucleotide targeting Arl4c (Arl4c ASO) that suppresses the proliferation of human hepatocellular carcinoma and colorectal cancer in the liver in vivo. Following these, in this study, we explored the clinical significance of high expression of Arl4c in patients with lung adenocarcinoma and examined whether Arl4c ASO could be an effective treatment for lung adenocarcinoma.
Arl4c expression was immunohistochemically examined in 161 patients who underwent surgical resection at Osaka University Hospital. Some lung adenocarcinomas progress gradually from precancerous atypical adenomatous hyperplasia (AAH) to carcinoma in situ (AIS), minimally invasive carcinoma (MIA), and invasive adenocarcinoma (IA). Higher Arl4c expression was detected more frequently in AAH than in adenocarcinoma patients. Among the lung adenocarcinoma patients without AAH lesions, high expression rate of Arl4c increased gradually as AIS (23.3%), MIA (36.3%) and IA (50.6%) as the histological type progressed. The Arl4c high expression patients tended to have high FDG accumulation on PET-CT, which is an indicator of lung cancer malignancy (p=0.01). In addition, the Arl4c high expression patients had a shorter relapse-free survival time than the low Arl4c expression patients (p=0.0128).
Furthermore, we noted that Arl4c was frequently expressed in AAH. To clarify the function of Arl4c in precancerous epithelial cells, we established an Arl4c overexpressing immortalized normal human airway epithelial cells (SAEC) and examined its phenotype. Arl4c expression in SAECs promoted cell proliferation. These results indicated Arl4c might enable the epithelial cells to abnormally proliferate in precancerous lesions.
Intratracheally administered Arl4c ASO effectively reduced Arl4c expression and suppressed the growth of orthotopic lung tumors consisting of both the Kras mutant cell line A549 and the EGFR mutant cell line H1975. Based on the present study results, Arl4c ASO may have therapeutic potential for Arl4c-expressing lung cancer with not only Kras- or EGFR-activating mutations.
In conclusion, Arl4c expression may play an important role in the carcinogenesis of lung cancer, and furthermore, the high expression of Arl4c may be a poor prognostic factor in lung adenocarcinoma. We also believe that ASO targeting Arl4c can be expected as a new treatment for lung cancer.
 GREB1, a novel target of Wnt signaling, promotes the development of hepatoblastoma
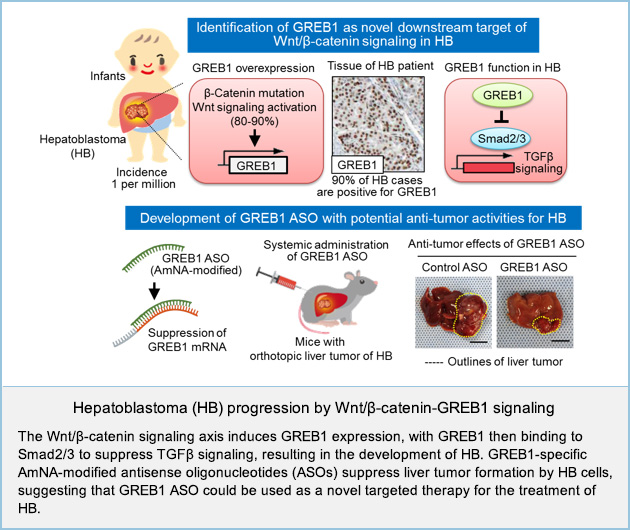
Hepatoblastoma (HB) is a rare form of liver cancer affecting just a few individuals per million. While advances in surgery and chemotherapy have meant that the prognosis for hepatoblastoma patients is generally quite good, aggressive forms of the disease leave some young patients with few treatment options and poor long-term survival rates. Although HB has highest association with mutational activation of β-catenin, the underlying mechanism by which Wnt/β-catenin signaling induces HB tumor formation is unknown.
Here we identify growth regulation by estrogen in breast cancer 1 (GREB1), which is a well-known estrogen-responsive gene implicated in the growth of breast cancer cells, as a previously unknown target gene of Wnt/β-catenin signaling. GREB1 was overexpressed in tumor lesions of 10/11 HB tissues (90.9%) and in public dataset of patients with HB. The region-specific expression pattern of GREB1 in HB tissues tended to be positively correlated with the accumulation of β-catenin.
GREB1 promoted proliferation and suppressed differentiation of HB cells. GREB1 bound Smad2/3 and competed with histone acetyl transferase p300 in the nucleus, resulting in the inhibition of TGFβ-dependent cytostasis.
Forced expression of & β-catenin, YAP, and c-Met (BYM) induced HB-like liver tumor in mice, with an increase in GREB1 expression and HB markers. GREB1 knockdown suppressed expression of HB marker genes and tumor formation in BYM mice. Furthermore, we synthesized an amido-bridged nucleic acid (AmNA)-modified antisense oligonucleotides (ASOs) that target human GREB1. These small single-stranded DNA molecules specifically interfere with the production of target proteins by binding to the mRNA. GREB1 depletion by ASOs injection suppressed the formation of hepatoblastoma tumors in mice.
These findings uncover a previously unrecognized involvement of Wnt/β-catenin-GREB1-Smads axis in HB pathogenesis, suggesting that GREB1 represents a potential therapeutic target for β-catenin-driven HB.
 Dynamic palmitoylation of DKK1 receptors, CKAP4 and LRP6, regulates DKK1 signaling activity
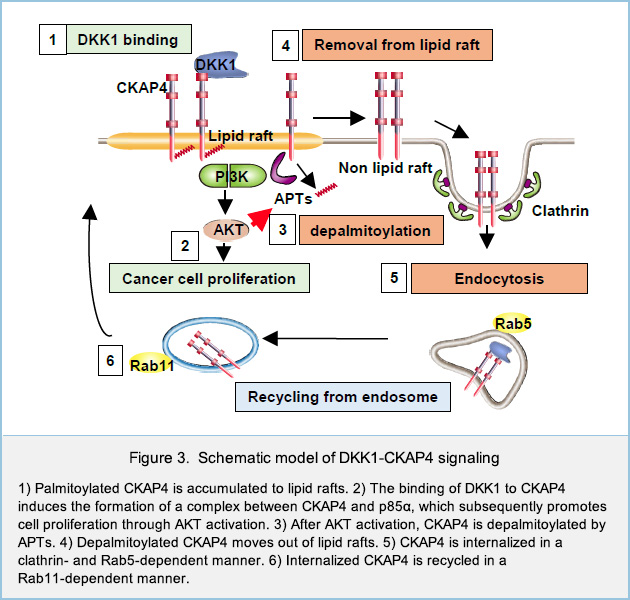
DKK1 is a secretory protein and antagonizes Wnt signaling by binding to the Wnt coreceptor low-density lipoprotein receptor-related proteins 6 (LRP6). Previously we identified CKAP4, a type II single transmembrane protein, as a novel Dickkopf1 (DKK1) receptor. DKK1-CKAP4 signaling activated the phosphatidylinositol 3-kinase (PI3K)-AKT pathway, resulting in cancer cell proliferation. CKAP4 has been shown to be modified with palmitate at Cys100, but its physiological roles in DKK1-CKAP4 signaling was not clarified. Besides, although LRP6 is palmitoylated, whether palmitoylation of LRP6 has some roles in DKK1 signaling remains to be clarified. In this study, we elucidated the molecular mechanisms that control palmitoylation of two DKK1 receptors, CKAP4 and LRP6, and physiological roles of dynamic palmitoylation in the regulation of DKK1 signaling.
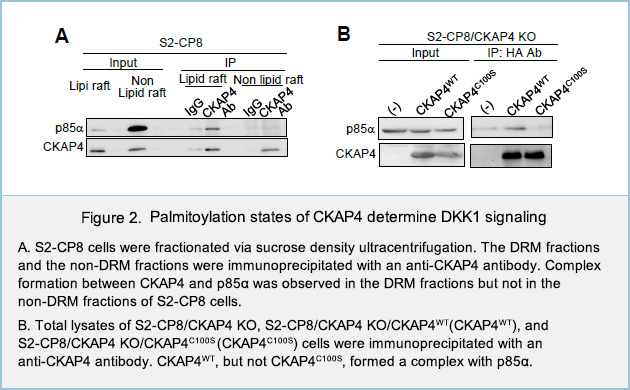
To investigate the roles of palmitoylation in both receptors, plasma membrane proteins of human pancreatic cancer cell line S2-CP8 cells were biotinylated and fractionated via sucrose density ultracentrifugation method. In the plasma membrane, most of WT CKAP4 proteins were detected in the detergent resistant membrane fractions (DRM), which reflect lipid rafts. On the other hand, palmitoylation-deficient CKAP4 mutant protein in which Cys100 was mutated to Ser (CKAP4C100S) were exclusively located at non-DRM fractions, which reflect non-lipid rafts (Fig. 1A).
In the Click chemistry assay, S2-CP8 cells which endogenous DKK1 was knocked out (S2-CP8/DKK1 KO) were metabolically labeled with the palmitate analogue 17-octadecynoic acid (17-ODYA), which was then modified with azide-biotin and used to precipitate palmitoylated proteins with NeutrAvidin-agarose beads. When S2-CP8/DKK1 KO cells were stimulated with recombinant DKK1, the palmitoylation levels of both CKAP4 and LRP6 at the plasma membrane of S2-CP8/DKK1 KO cells were reduced (Fig. 1B). Interestingly, at the same assay condition, sucrose density ultracentrifugation method revealed that both CKAP4 and LRP6, which located in lipid rafts at steady-state conditions, were moved to non-lipid rafts in response to DKK1 (Fig. 1C). Taken together with other results, these results suggested that two DKK1 receptors, CKAP4 and LRP6, are localized to the lipid rafts in the palmitoylation-dependent manner, and that DKK1 induces the depalmitoylation of both receptors, followed by removal of both receptors from lipid rafts.
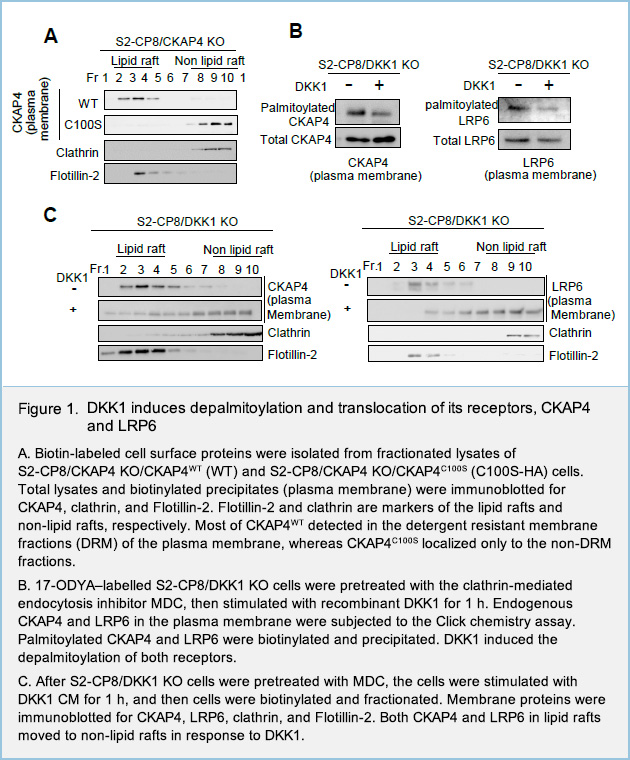
We next examined the role of localization of CKAP4 to lipid rafts in DKK1-CKAP4 signaling. DKK1 promoted the formation of a complex between CKAP4 and the p85α regulatory subunit of PI3K. Complex formation between CKAP4 and p85α was observed in the DRM fractions but not in the non-DRM fractions of S2-CP8cells (Fig. 2A). Moreover, CKAP4WT, but not CKAP4C100S, formed a complex with p85α when expressed in S2-CP8/CKAP4 KO cells (Fig. 2B), indicating that CKAP4 interacts with PI3K in lipid rafts. Thus, DKK1 induces depalmitoylation of CKAP4, resulting in its translocation from lipid rafts to non-lipid rafts, and that palmitoylation of CKAP4 is critical for regulating DKK1 signaling (Fig. 3). LRP6 is also depalmitoylated in response to DKK1, and moved from lipid rafts to non-lipid rafts.
DKK-CKAP4 signaling promotes tumorigenesis in pancreas, lung, and esophageal cancers
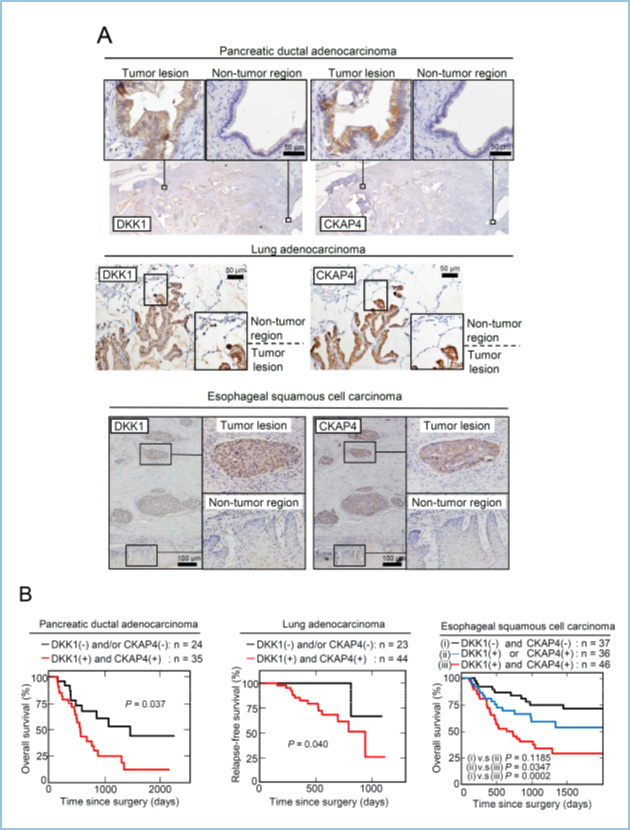
Dickkopf 1 (DKK1) is a secretory protein and antagonizes oncogenic Wnt signaling by binding to the Wnt coreceptor low-density lipoprotein receptor-related proteins 6 (LRP6). DKK1 is also suggested to regulate its own signaling to promote cancer cell proliferation, but the mechanism has not been understood. We screened DKK1-binding proteins and identified cytoskeleton-associated protein 4 (CKAP4) as a novel DKK1 receptor. DKK1-CKAP4 signaling activated AKT through the formation of a complex between CKAP4 and phosphatidylinositol 3-kinase (PI3K), resulting in cancer cell proliferation.
In pancreatic ductal adenocarcinoma (PDAC), DKK1 and CKAP4 were strongly detected in 45/59 (76.3%) and 39/59 (66.1%) cases, respectively. In lung adenocarcinoma, 53/67 (79.1%) and 50/67 (74.6%) cases were positive for DKK1 and CKAP4 expression, respectively. In lung squamous cell carcinoma (SCC) cases, DKK1 and CKAP4 were positive in 45/61 (73.8%) and 42/61 (68.9%) cases, respectively. In esophageal SCC (ESCC) cases, DKK1 and CKAP4 were strongly expressed in 66/119 (55.5%) and 62/119 (52.1%) cases, respectively. Both proteins were minimally detected in non-tumor regions under our staining conditions. In PDAC and ESCC cases, overall survival was significantly reduced in both DKK1 and CKAP4 positive patients compared with DKK1 and/or CKAP4 negative patients. In lung cancer cases, both DKK1 and CKAP4 expression was negatively correlated with relapse-free survival. These indicate that their simultaneous expression is negatively correlated with prognosis and relapse-free survival of pancreatic and lung cancers.
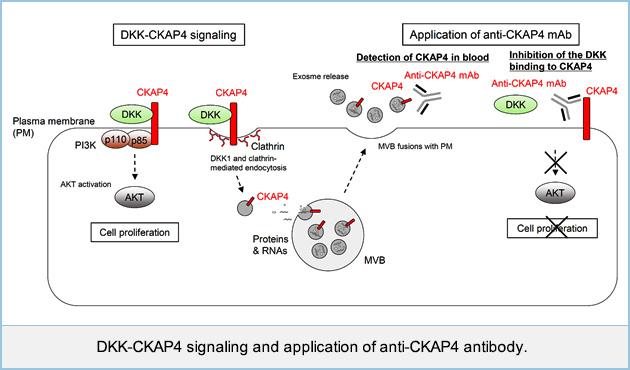
The role of DKK1-CKAP4 signaling in tumorigenesis in vivo was investigated by subcutaneously implanting cancer cells into the flanks of immunodeficient mice. The volumes and weights of the xenograft tumors derived from S2-CP8 pancreatic, A549 lung, and TE-8 esophageal cancer cells stably expressing CKAP4 or DKK1 shRNA were less than those of control tumors. The rabbit polyclonal anti-CKAP4 antibody blocked the binding of DKK1 to CKAP4, thereby suppressing AKT activity in cancer cells and xenograft tumor formation in immunodeficient mice. Thus, CKAP4 might represent a novel therapeutic target for cancers expressing DKK1 and CKAP4. We are currently generating monoclonal antibodies that specifically block DKK1-CKAP4 signaling and would like to develop a novel cancer therapy.
In addition to DKK1, other DKK family members also bound to CKAP4 and increased cellular proliferation. Particularly, DKK3 exhibits structural divergence from other family members; and its corresponding receptors remain to be identified. DKK3 was expressed in tumor lesions of esophageal squamous cell carcinoma (ESCC) cases; simultaneous expression of DKK3 and CKAP4 was associated with poor prognosis. p63, a p53-related transcriptional factor frequently amplified in ESCC, and bound to the upstream region of the DKK3 gene. Knockdown of p63 decreased DKK3 expression in ESCC cells. Expression of p63 and DKK3 increased the size of tumor-like esophageal organoids, and an anti-CKAP4 antibody inhibited growth of esophageal organoids. Taken together, these results suggest that the DKK3-CKAP4 axis might also serve as a novel molecular target for ESCC.
We generated anti-CKAP4 monoclonal antibodies(mAbs). Anti-CKAP4 mAbs suppressed xenograft tumor formation in immunodeficient mice and extended the survival of mice receiving intraperitoneal or orthotopic injection of pancreatic cancer cells. We found that CKAP4 is released with exosomes from pancreatic cancer cells, in which DKK1 was overexpressed and CKAP4 was localized to the plasma membrane, and that the secretion of CKAP4-containing exosomes is mediated by DKK1-dependent endocytosis. We developed enzyme-linked immuno-sorbent assay (ELISA) to detect CKAP4 in pancreatic cancer patient sera using anti-CKAP4 mAbs. The serum CKAP4 levels were higher in pancreatic cancer patients than healthy individuals. CKAP4 secreted in exosomes may represent a biomarker for pancreatic cancer. Furthermore, anti-CKAP4 mAbs can contribute to the development of novel diagnostic methods and therapeutics.
 Development of antisense oligonucleotide against ARL4C for liver tumor
ADP-ribosylation factor-like 4c (ARL4C), a member of ARL family proteins, is a target protein that is expressed by the activation of Wnt/β-catenin and EGF/RAS/ mitogen-activated protein kinase (MAPK) signaling and plays an important role in both epithelial morphogenesis and tumorigenesis.
ARL4C expression was analyzed in primary hepatocellular carcinoma (HCC) using immunohistochemistry. Among the 128 HCC samples, ARL4C was detected and positive in 33 tumors (25.8%) but only minimally detected in matched non-tumor regions. Furthermore, relapse-free survival was significantly decreased in patients with ARL4C-positive tumors compared to patients with ARL4C-negative tumors in HCC (P = 0.033).
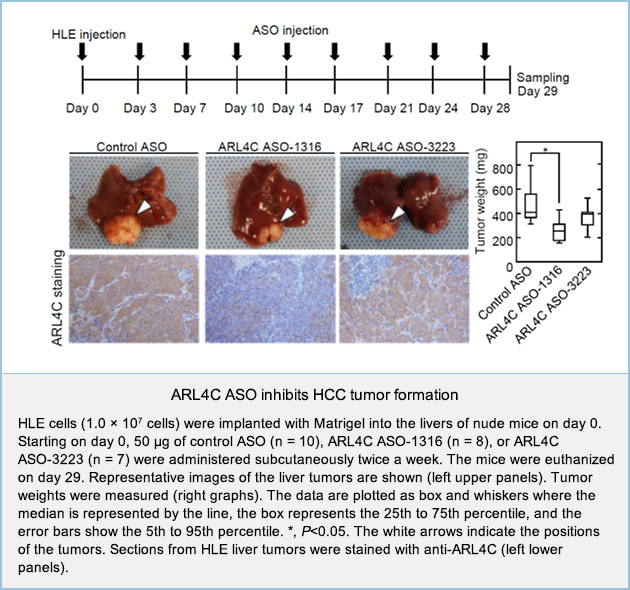
To clarify whether ARL4C could be a therapeutic target for the treatment of liver tumors, ARL4C antisense oligonucleotides (ASOs) were generated and their effects on cancer cells were examined. Among several ARL4C ASO candidates, ARL4C ASO-1316 and ARL4C ASO-3223 were selected based on their strong downregulation of ARL4C mRNA expression in HLE HCC cells at low doses. ARL4C ASO-1316 and ASO-3223 suppressed the migration of HLE cells, which was rescued by the overexpression of ARL4C, excluding off-target effects by the ARL4C ASOs. ARL4C ASOs also inhibited HLE cell proliferation.
HLE cells were injected into the liver, and the mice were subcutaneously injected with control ASO and two ARL4C ASOs. ARL4C ASO-1316, but not ASO-3223 suppressed HLE tumor formation in the liver and ARL4C expression in HLE cells compared to control ASO (Figure 1). In addition, ARL4C ASO-1316 was also effective for metastatic liver tumors induced by HCT116 colon cancer cells.
In conclusion, ARL4C is involved in liver cancer cell proliferation and ARL4C ASO represents the novel treatment for primary and metastatic liver cancers.
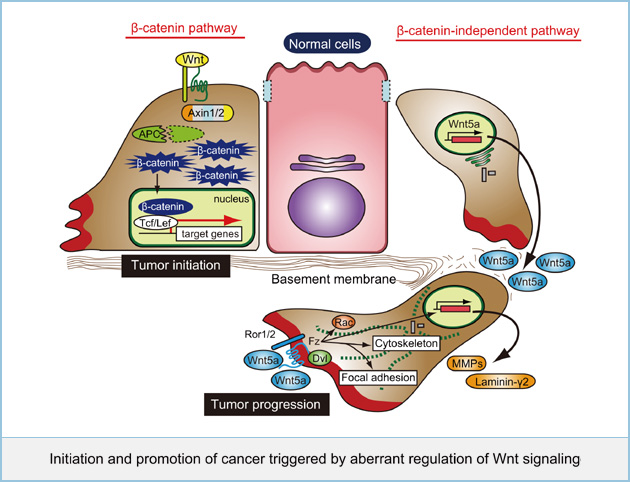
 Initiation and promotion of cancer triggered by aberrant regulation of Wnt signaling
Mutations of genes involved in Wnt signaling have been reported to associate with many cancers. In particular, mutations of core components in the β-catenin pathway such as APC, β-catenin, and Axin have been well known to lead to aberrant activation of this pathway, resulting in various cancers, including colorectal and hepatocellular cancers. Recently several groups including my lab have reported that abnormalities of the β-catenin-independent pathway also associate with progression and aggressiveness in several cancers, including melanoma, lung, gastric, and prostate cancers. The β-catenin-independent pathway has several activities to regulate cytoskeletal rearrangements, cell polarity, and intracellular calcium-dependent signaling to stimulate cell migration and adhesion. However, the molecular mechanism how aberrant regulation of its signaling pathway contributes to cancer progression remains unclear. We are currently studying the molecular actions of Wnt5a, a representative ligand for the β-catenin-independent pathway, to lead to initiation and promotion of cancer.
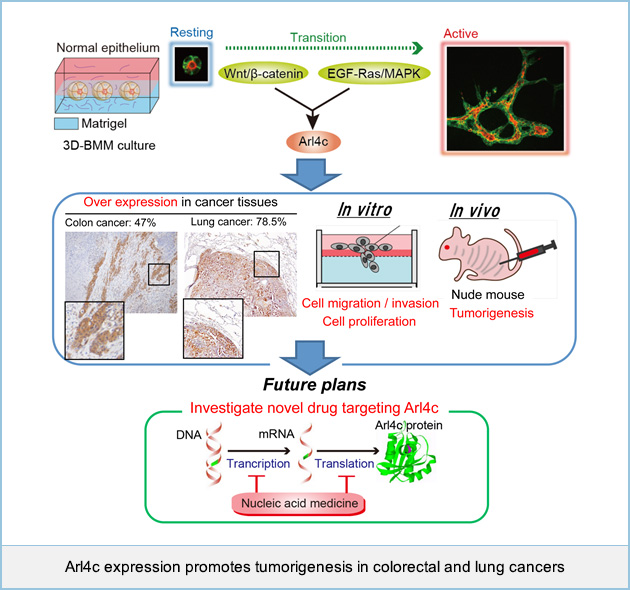
 Arl4c expression promotes tumorigenesis in colorectal and lung cancers
Wnt/β-catenin and EGF/Ras signaling have been reported to associate with many human cancers, including colorectal and lung cancers. We recently found that ADP-ribosylation factor (ARF)-like 4c (Arl4c) expression induced by a combination of Wnt/β-catenin and EGF/Ras signaling in normal epithelial cells grown in three-dimensional culture promotes cellular migration and proliferation, resulting in formation of tubular structures. Although the process of epithelial morphogenesis resembles tumor formation in that both expand in the mensenchyme. The involvement of Arl4c in tumorigenesis has remained unclear. Immunohistochemical analysis showed that Arl4c was not observed in non-tumor regions, but was overexpressed in tumor lesions in 50 ~ 80% of colorectal and lung cancers. In several colorectal and lung cancer cell lines, Arl4c expression was upregulated through Wnt/β-catenin or Ras/MAPK signaling. Inhibition of Arl4c expression in HCT116 colorectal cancer cells and A549 lung adenocarcinoma cells reduced migration, invasion, and proliferation capabilities both in vitro and in vivo. Furthermore, direct injection of Arl4c siRNA into HCT116 cell-derived tumors (in vivo treatment with siRNA) inhibited tumor growth in immunodeficient mice. These findings suggested that Arl4c could be a novel marker for cancers and a molecular target for cancer therapy. We are currently developing new nucleic acid based medicine representing therapeutic target for cancers with Arl4c overexpression.
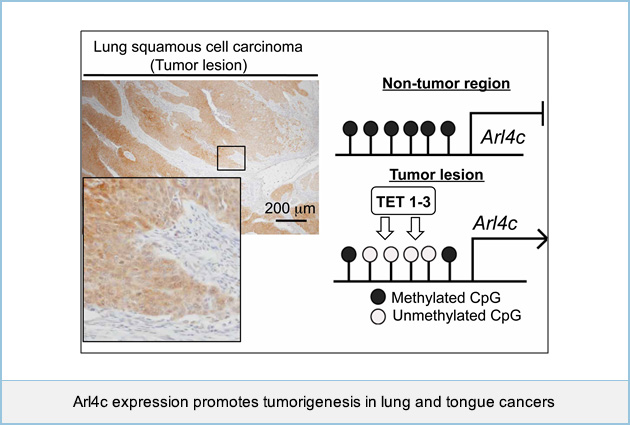
 Epigenetic upregulation of Arl4c expression, due to DNA hypomethylation in the 3'-untranslated region, in lung squamous cell carcinoma
We recently found that expression of ADP-ribosylation factor (ARF)-like 4c (Arl4c) was highly expressed in tumor lesions of colon and lung adenocarcinomas, where genetic alterations of Wnt/β-catenin and EGF/Ras pathways are frequently observed, and Arl4c expression promoted migration, invasion, and proliferation of cancer cells both in vitro and in vivo. However, it is unclear whether Arl4c is overexpressed in tumors where aberrant activation of growth factor signaling, such as Wnt/β-catenin or EGF/Ras pathways, does not occur. Immunohistochemical analysis showed that Arl4c was not observed in non-tumor regions, but was overexpressed in tumor lesions in 70 ~ 80% of lung and tongue squamous cell carcinoma (SCC). Knockout of Arl4c expression in NCI-H520 lung SCC cells and SAS tongue SCC cells reduced proliferation and migration capabilities in vitro. Although inhibition of Wnt/β-catenin or Ras/MAP kinase signaling did not decrease Arl4c expression in NCI-H520 cells, Arl4c DNA was clearly hypomethylated in the 3'-untranslated region (UTR). Ten-eleven translocation methylcytosine dioxygenase (TET) enzyme, which mediates DNA demethylation, was highly expressed in NCI-H520 cells. Knockout of TET family proteins (TET1-3) in NCI-H520 cells promoted DNA methylation in the 3'-UTR, leading to the decrease in Arl4c expression. The analyses with The Cancer Genome Atlas (TCGA) dataset (379 lung cancer cases) revealed that DNA methylation in the 3'-UTR of Arl4c gene was lower than in non-tumor regions in Arl4c-positive lung SCC. The results were consistent with our observations. These findings suggested that Arl4c contributes to oncogenesis as a result of its overexpression by multiple mechanisms and a molecular target for cancer therapy. We are currently developing new nucleic acid based medicine representing therapeutic target for cancers with Arl4c overexpression.
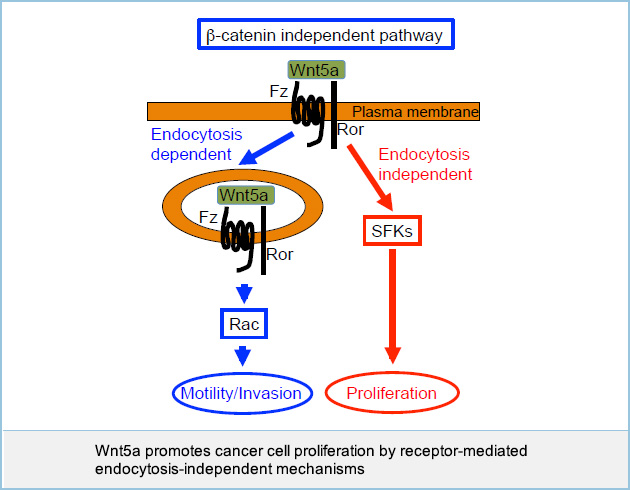
 Wnt5a promotes cancer cell proliferation by receptor-mediated endocytosis-independent mechanisms
Wnt5a expression was previously believed to be associated with cancer cell motility but not proliferation. For instance, Wnt5a promoted invasion but not proliferation in some cancer cells such as KKLS gastric cancer cells. It was also reported that Wnt5a is also implicated in cancer cell proliferation, but the mechanism was not clear. Wnt5a indeed stimulated both invasion and proliferation of certain types of cancer cells, including HeLaS3 cervical cancer cells and A549 lung cancer cells.
Although an anti-Wnt5a polyclonal neutralizing antibody was generated previously, the neutralizing activity of polyclonal antibodies varies among antibody lots. Therefore, the generation of a neutralizing monoclonal antibody against Wnt5a was necessary to control quality and quantity of the antibody. We generated an anti-Wnt5a monoclonal antibody by using a phage library. An anti-Wnt5a monoclonal antibody inhibited invasion but not proliferation of HeLaS3 and A549 cells. Wnt5a activated Src family kinases (SFKs) and Wnt5a-dependent cancer cell proliferation was dependent on SFKs, yet blockade of receptor-mediated endocytosis did not affect cancer cell proliferation and SFK activity. These results suggest that Wnt5a promotes invasion and proliferation of certain types of cancer cells through receptor-mediated endocytosis-dependent and -independent mechanisms, respectively.
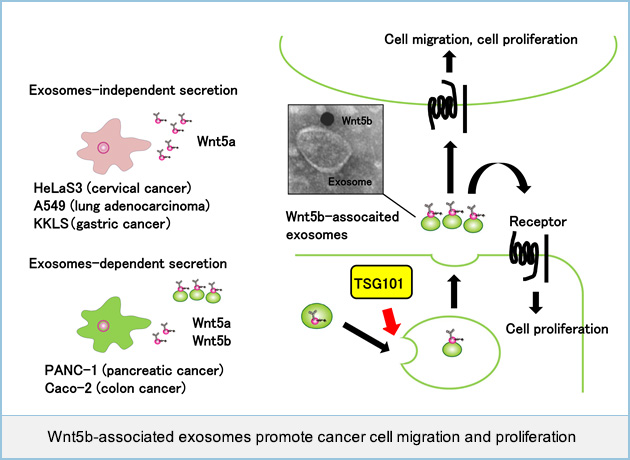
 Cancer cells secrete Wnt5b-associated exosomes
Wnt5b, a member of the same family of proteins as Wnt5a of which overexpression is associated with cancer aggressiveness, is suggested to be involved in cancer progression, however the details remain unclear. It was reported that some Wnts were secreted with extracellular microvesicles, such as exosomes, which were recovered in the 100,000 × g centrifugation precipitation fractions from conditioned medium (CM). Wnt5b released from PANC-1 pancreatic cancer cells or Caco-2 colon cancer cells transiently expressing Wnt5b, was recovered in precipitation fractions. However, some cancer cell lines, including HeLaS3 cervical cancer, A549 lung adenocarcinoma, and KKLS gastric cancer, expressed Wnt5a endogenously, and most Wnt5a secreted in CM was present in supernatant fractions.
To examine whether Wnt5b is indeed associated with exosomes, we generated a plasmid vector expressing hemagglutinin (HA)-tagged Wnt5b (HA-Wnt5b), by introducing HA tag sequence at position Arg36, located in a weakly conserved surface region opposite the Frizzled receptor binding site. HA-Wnt5b was recovered in precipitation fractions from CM of PANC-1 cells expressing HA-Wnt5b. Immunoelectron microscopy revealed that HA-Wnt5b is present on exosomes isolated from PANC-1 cells expressing HA-Wnt5b.
To examine endogenous Wnt5b activity in a PANC-1 cell, the WNT5B gene was successfully knocked out by a CRISPR/Cas9 system. Precipitation fractions from CM of PANC-1 cells showed Wnt5b activity, but those from WNT5B knockout (KO) PANC-1 cells dramatically decreased the activity. Exosomes isolated from wild-type PANC-1 cells, but not those from WNT5B KO PANC-1 cells, stimulated cell migration of and increased 5-ethynl-2'-deoxyuridine (EdU) incorporation into A549 cells. In Caco-2 cells expressing Wnt5b, knockdown of TGS101, which is a maker of exosomes, suppressed the secretion of Wnt5b-associated exosomes and inhibited Wnt5b-dependent cell proliferation.
Taken together, these results suggested that Wnt5b-associated exosomes promote cancer cell migration and proliferation in a paracrine manner.
 Development of diagnostic and therapeutic devises for Wnt5a signaling as a molecular target in cancer
We found that Wnt5a is involved in malignancy of some kind of human cancer including gastric and prostate cancers. Immunohistochmical studies showed that expression of Wnt5a is observed in about 30% of gastric and prostate cancer cases. Expression of Wnt5a was associated with aggressiveness and poor prognosis of gastric cancer. In addition, injection of polyclonal anti-Wnt5a antibody suppressed the frequency of tumor metastasis of gastric cancer cells. Although these findings suggested that Wnt5a could be a novel target for cancer therapy, there are no specific therapeutic devises for Wnt5a-mediated cancers. To identify specific therapeutic devises for Wnt5a signaling as a molecular target in cancer, we are trying to produce neutralizing monoclonal antibodies to Wnt5a. Furthermore, we are going to identify small molecules inhibiting the secretion of Wnt5a or the binding of Wnt5a to its receptors by using a high-throughput screening (HTS).
 2. Epithelial morphogenesis regulated by growth factor signaling
Regulation of fetal lung epithelial morphogenesis
During the later stages of lung development, two types of pneumocytes, cuboidal type II (AECII) and flattened type I (AECI) alveolar epithelial cells, form distal lung saccules. We highlighted how fibroblasts-expressing Mark1 is required for distal lung sacculation. In Mark1 knockout (KO) mice, distal sacculation and AECI flattening were significantly impaired.
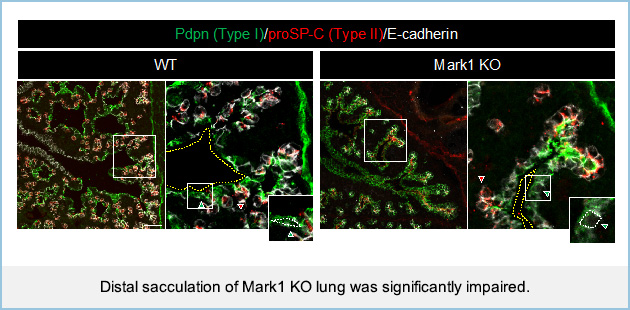
Fetal epithelial cells generated alveolar organoids and were differentiated into pneumocytes when cocultured with fibroblasts. However, the size of organoids decreased and AECI flattening was impaired in the presence of Mark1 KO fibroblasts. In Mark1 KO fibroblasts themselves, cilia formation and the Hedgehog pathway were suppressed, resulting in the loss of type I collagen expression. The addition of type I collagen restored AECI flattening in organoids-cocultured with Mark1 KO fibroblasts and rescued the decreased size of organoids. Mathematical modeling of distal lung sacculation supported the view that AECI flattening is necessary for properly formed saccule-like structures. These results suggest that Mark1-mediated fibroblast activation induces AECI flattening, thereby regulating distal lung sacculation.
Roles of Wnt signaling in fetal lung epithelial morphogenesis
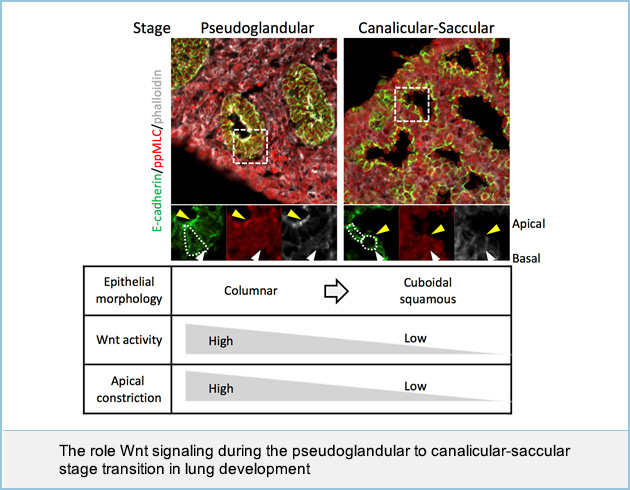
In lung development, the apically constricted columnar epithelium forms numerous buds during the pseudoglandular stage. Subsequently, these epithelial cells change shape into the flat or cuboidal pneumocytes that form the air sacs during the canalicular and saccular (canalicular-saccular) stages, yet the impact of cell shape on tissue morphogenesis remains unclear. Here, we show that the expression of Wnt components is decreased in the canalicular-saccular stages, and that genetically constitutive activation of Wnt signaling impairs air sac formation by inducing apical constriction in the epithelium as seen in the pseudoglandular stage. Organ culture models also demonstrate that Wnt signaling induces apical constriction through apical actomyosin cytoskeletal organization. Mathematical modeling reveals that apical constriction induces bud formation and that loss of apical constriction is required for the formation of an air sac-like structure. We identify MAP/microtubule affinity-regulating kinase 1 (Mark1) as a downstream molecule of Wnt signaling and show that it is required for apical cytoskeletal organization and bud formation. These results suggest that Wnt signaling is required for bud formation by inducing apical constriction during the pseudoglandular stage, whereas loss of Wnt signaling is necessary for air sac formation in the canalicular-saccular stages.
 Regulation of epithelial morphogenesis by soluble growth factors and adhesion
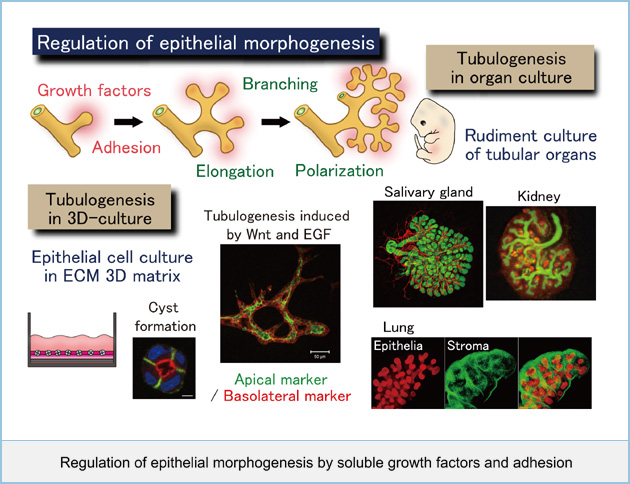
It is important to understand how individual cells organize into multicellular tissues. Extracellular matrix and growth factors are essential for establishing and maintaining epithelial tube formation by controlling elongation, polarization, and branching. We are interested in how multicellular cell motility, polarity, proliferation, and differentiation are coordinately controlled at molecular and cellular level to form epithelial tubules. To address these questions, we are currently examining the mechanism of polarized cyst formation and analyzing the roles of soluble factors such as Wnt and EGF in branching morphogenesis by the use of three-dimensional culture of epithelial cells in vitro. In addition, using organs from embryo such as salivary and mammary glands, kidney, and lung, we will clarify 1) the mechanisms for collective cell elongation, polarization, and branching induced by specific growth factors and adhesion in each organ and 2) the common or different mechanisms in epithelial tube formation from various tissues.
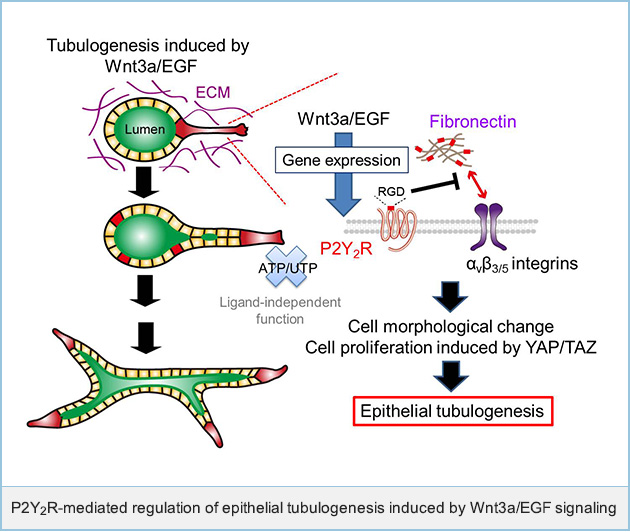
 Regulation of tubular formation of IEC6 cells by P2Y2 receptor expression
We developed a new in vitro tubular formation assay using rat intestinal epithelial cells (IEC6 cells) in 3D Matrigel. IEC6 cells formed cysts in 3D Matrigel, and simultaneous stimulation with Wnt3a and EGF (Wnt3a/EGF) induced tubular formation. Using DNA microarray analyses, we have found that Wnt3a/EGF induces the expression of ADP-ribosylation factor (Arf)-like 4c (Arl4c) and Arl4c promotes tubular formation through the regulation of Rac and Rho activities. In this study, DNA microarray analyses also revealed that P2ry2 (P2Y2receptor (P2Y2R) gene) mRNA is expressed by Wnt3a/EGF during tube formation.
P2Y2R is a nucleotide-activated G-protein coupled receptor and binds to both purine and pyrimidine, ATP and UTP. Although P2Y2R regulates various cellular functions, for example, cotransmission and neuromodulation in nerve cells, the involvement of P2Y2R expression in epithelial tubulogenesis has not been previously studied. Then, we tried to clarify P2Y2R-mediated regulation of tubulogenesis induced by Wnt3a/EGF.
We examined the involvement of P2Y2R in epithelial tubulogenesis induced by Wnt3a/EGF. Knockdown of P2Y2R using siRNA suppressed Wnt3a/EGF-induced tubular formation, and P2Y2R-HA expression rescued the phenotypes. These results indicate that P2Y2R is required for tubular formation induced by Wnt3a/EGF.
We examined the involvement of P2Y2R ligands in epithelial tubulogenesis. A P2Y2R mutant that lacks nucleotide responsiveness rescued the phenotypes resulting from P2Y2R knockdown, suggesting that nucleotide-dependent responses are not required for P2Y2R functions in tubular formation.
To clarify ligand-independent functions of P2Y2R in tubulogenesis, we focused on the Arg-Gly-Asp (RGD) sequence of P2Y2R which has been shown to associate with integrins. A P2Y2R mutant that lacks the integrin-binding activity lost the ability to regulate tubular formation. These results indicate that binding of P2Y2R to integrins is required for tubular formation induced by Wnt3a/EGF.
Additionally, P2Y2R expression inhibited the formation of a complex between fibronectin which contains RGD sequences, and integrins. Appropriate inhibition of the interaction between fibronectin and integrins by the cyclic RGD peptide (RGDfV) and fibronectin siRNA enhanced, but a large amount of fibronectin suppressed Wnt3a/EGF-induced tubular formation.
The present study demonstrated P2Y2R expression induced by Wnt3a/EGF causes tubular formation of IEC6 cells by preventing the binding of integrins and fibronectin rather than mediating nucleotide responses.
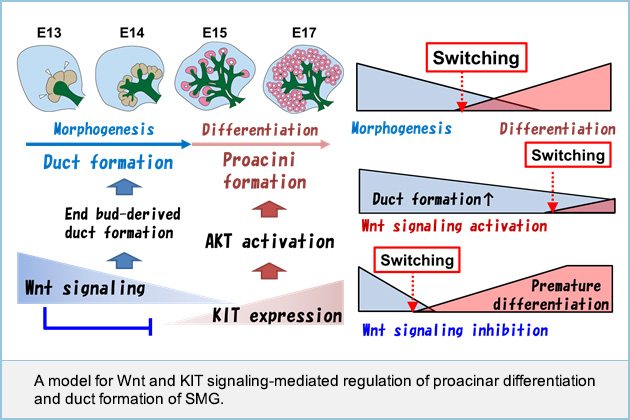
 Regulation of developmental switching between "morphogenesis" and "differentiation"
Growth factor signaling is involved in the development of various tubular organs, but how signaling regulates organ early morphogenesis and late differentiation remains to be clarified. Here, we show how Wnt signaling controls epithelial tubular branching morphogenesis and functional differentiation using the salivary gland as a model. Experiments using genetically manipulated mice and organ cultures revealed that Wnt signaling at early stage (E12-E14) of submandibular salivary gland (SMG) development inhibits end bud morphogenesis and differentiation into proacini by suppressing KIT expression, and concomitantly increased the expression of distal progenitor markers such as Sox10 and KRT14. In addition, Wnt signaling at the SMG development early stage promoted end bud cell proliferation, leading to duct formation. In contrast, Wnt signaling loss at a late stage (E15-E18) of SMG development promoted end bud differentiation into proacini and suppressed duct formation. Our results suggested that Wnt signaling fine-tunes the timing of end bud morphogenesis and differentiation into proacini, in co-operation with KIT signaling.
 3. Novel Functions of CKAP4
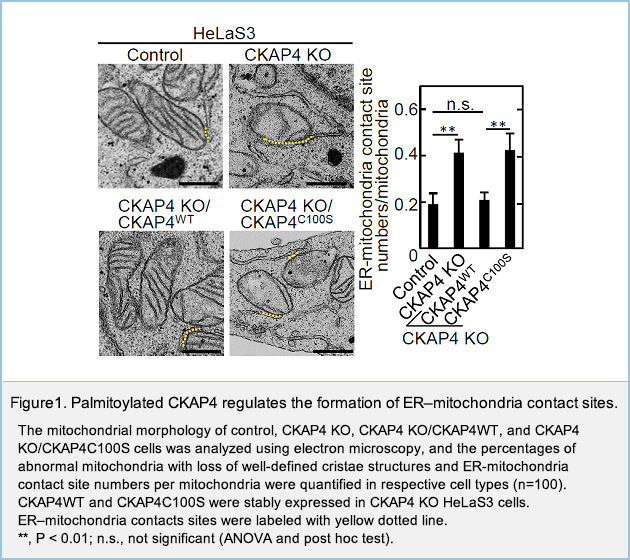
Palmitoylated CKAP4 regulates mitochondrial functions through an interaction with VDAC2 at ER–mitochondria contact sitesNew
Cytoskeleton-associated protein 4 (CKAP4) is palmitoylated type II transmembrane protein localized to the endoplasmic reticulum (ER). Although CKAP4 is likely to be involved in the regulation of the morphology and functions of the ER, the roles of CKAP4 in the functions of other organelles remain obscure. We found the knockout (KO) of CKAP4 in HeLaS3 cells induced the alterations of mitochondrial structures and increased the number of ER-mitochondria contact sites, in addition to ER morphological changes.
To understand the involvement of CKAP4 in mitochondrial functions, the binding proteins of CKAP4 were explored, enabling identification of the mitochondrial porin voltage-dependent anion-selective channel protein 2 (VDAC2), which is localized to the outer mitochondrial membrane. Palmitoylation at Cys100 of CKAP4 was required for the binding of CKAP4 and VDAC2. In CKAP4 KO cells, the binding of inositol trisphosphate receptor (IP3R) and VDAC2 was enhanced, the intramitochondrial Ca2+ concentration increased, and the mitochondrial membrane potential decreased. In addition, CKAP4 KO decreased the oxidative consumption rate, in vitro cancer cell proliferation under low-glucose conditions, and in vivo xenograft tumor formation. The phenotypes were not rescued by a palmitoylation-deficient CKAP4 mutant.
These results suggest that CKAP4 plays a role in maintaining mitochondrial functions through the binding to VDAC2 at ER–mitochondria contact sites and that palmitoylation is required for this novel function of CKAP4.
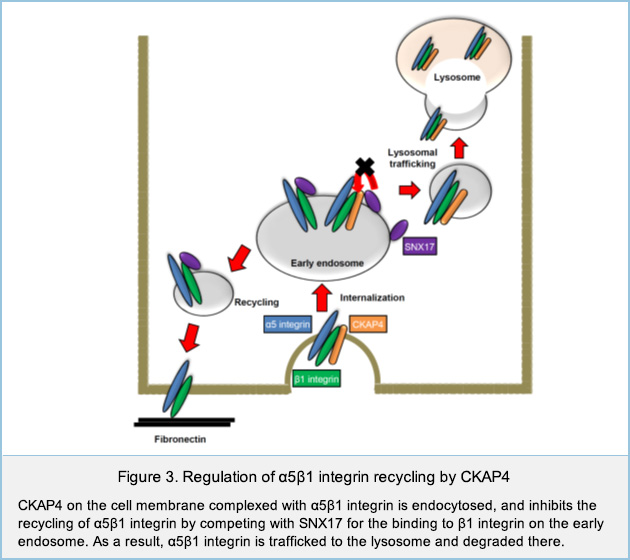
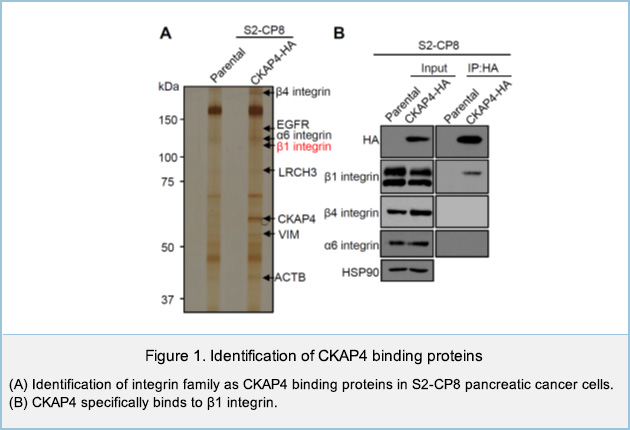
 Regulation of cell migration by cell surface CKAP4
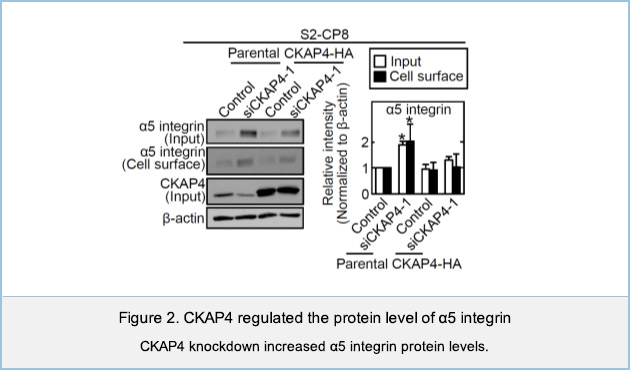
We have identified Cytoskeleton-associated protein 4 (CKAP4) as a novel receptor for Dickkopf1 (DKK1), a regulator of Wnt / β-catenin signaling. DKK1-CKAP4 signaling promoted cell proliferation through the phosphoinositide 3-kinase (PI3K)–AKT pathway in cancer cells. CKAP4 and DKK1 also stimulated cell migration, but the mechanism was unclear. Knockdown of CKAP4 on the cell surface enlarged the size of cell adhesion sites and inhibited cell migration independently of DKK1. In addition, CKAP4 bound to β1 integrin (Figure 1).
CKAP4 knockdown increased the protein level of α5 integrin, which is a partner of β1 integrin (Figure 2). CKAP4 on the cell membrane bound to β1 integrin and competed with SNX17 for the binding of to β1 integrin on the early endosome, thereby suppressing recycling of α5β1 integrin and promoting the traffic to the lysosome and degradation of CKAP4. Taken together, these results suggest that cell surface CKAP4 stimulates cell migration by decreasing the recycling of α5β1 integrin independently of DKK1 signaling (Figure 3).
4. Cellular functions regulated by Wnt signaling
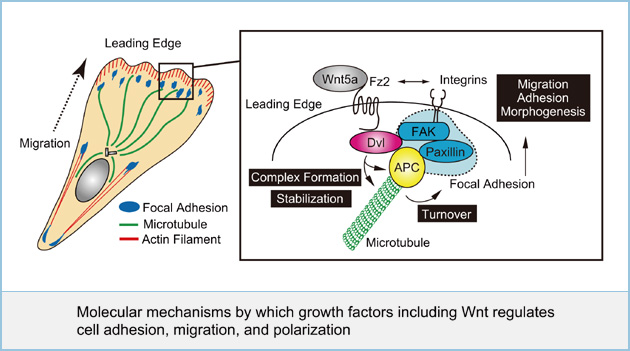
Molecular mechanisms by which growth factors including Wnt regulates cell adhesion, migration, and polarization
Cell polarization and the adhesion of a cell to a substrate are necessary for the cell to spread and migrate. When cells migrate, small focal adhesions, referred to as focal complexes at the cell periphery are formed. Upon the generation of tension, these focal complexes mature into large, organized adhesions, such as focal adhesions. These processes are regulated by small GTP-binding proteins Rac, Rho, and Cdc42. Although some growth factors such as EGF, VEGF, FGF, and Wnt regulate cell polarization, adhesion and migration, the molecular mechanism by which individual growth factors regulate these processes is poorly understood. We found that Wnt5a regulates cell migration and adhesion through the activation of focal adhesion kinase (FAK), a component of focal adhesion and Rac. Furthermore, we showed that dishevelled (Dvl), an essential component of Wnt signaling binds to adenomatous polyposis coli (APC), as microtubule (MT)-plus-end-binding protein at the leading edge of migrating cells, and both proteins associated with focal adhesion components. The binding of APC to Dvl is involved in stabilization of MTs at the cell periphery and Wnt5a-dependent focal adhesion turnover. We are currently analyzing the molecular mechanisms how other growth factors coordinately regulate cell adhesion, migration, and polarization.
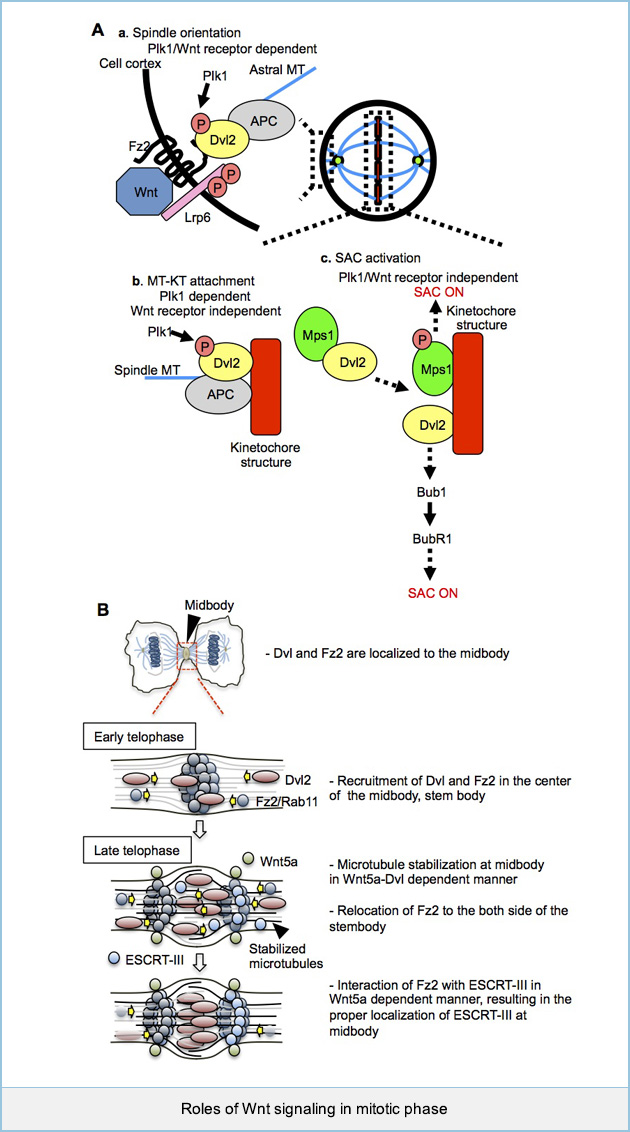
 Roles of Wnt signaling in mitotic phases
Wnt signaling is known to promote G1/S progression through the stimulation of gene expression, but whether this signaling regulates mitotic progression is not clear.
(A) We found that Dvls phosphorylated by Polo-like kinase 1 (Plk1) forms a complex with APC and Wnt receptors, Fz and LRP6 and that their interaction is required for spindle orientation (a). Phosphorylation of Dvl2 by Plk1 is also involved in the microtubule (MT) kinetochore interaction. On the other hands, Wnt receptors are not required in this process (b). Dvl2 activates spindle assembly checkpoint in Plk1-mediated Dvl2 phosphorylation-dependent and Wnt-receptors-independent manner. Furthermore, Dvl2 is involved in the activation of Mps1 and recruitment of Bub1 and BubR1 at kinetochore (c). These results suggest that Dvl2 is involved in mitotic progression by regulating the dynamics of MT plus-ends and the spindle assembly checkpoint (SAC) in Plk1-dependent and -independent manners.
(B) In addition to the involvement of Dvl2 in the early mitotic phase via the β-catenin pathway, Dvl2 is localized to midbody in late mitotic phase. Through our observations, we proposed a model that 1) at early telophase, Dvl2 and Fz2 are recruited to the center of midbody, stembody, and Wnt5a signaling is involved in the recruitment of Dvl2 to the dark zone, which depends on microtubules. 2) Accumulated Fz2 and Dvl2 at stembody enhance the stabilization of midbody microtubules, which also contributes to their efficient recruitment to the midbody. 3) At late telophase Fz2 relocates to both sides of the dark zone. Wnt5a signal strengthens the binding of Fz2 and CHMP4B and stabilizes the proper localization of CHMP4B to both sides of stembody, leading to complete abscission. These findings suggest that both β-catenin-dependent and -independent pathways are required for progression of mitosis in temporal and spatial restricted manner.
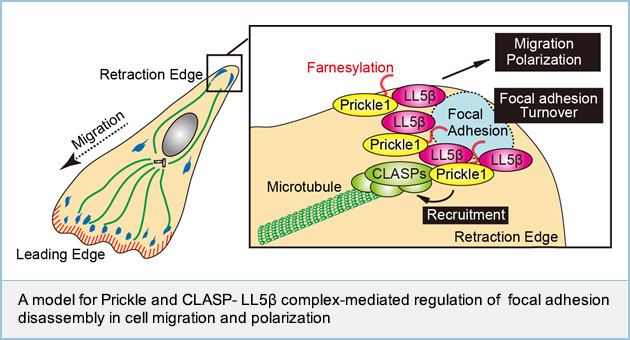
 Prickle1 promotes focal adhesion disassembly with the CLASP-LL5β complex
Prickle is known to play roles in planar cell polarity including convergent extension and cell migration; however, the detailed mechanism by which Prickle regulates cellular functions is not well understood. Here, we show that Prickle1 regulates front-rear polarization and migration of gastric cancer MKN1 cells. Prickle1 preferentially accumulated at the cell retraction site in close proximity to paxillin at focal adhesions (FAs). Prickle1 dynamics were associated with those of paxillin during FA disassembly. Furthermore, Prickle1 was required for FA disassembly. CLASP and LL5β have been reported to form a complex surrounding focal adhesions and controls microtubule-dependent FA disassembly. Prickle1 bound to CLASP and LL5β, and was required for the LL5β-dependent accumulation of CLASP at the cell edge. Knockdown of CLASP and LL5β suppressed Prickle1-dependent cell polarization and migration. Prickle1 localized to the membrane through its C-terminal farnesyl moiety, and the membrane localization was necessary for Prickle1's ability to regulate migration, bind to CLASP and LL5β, and promote microtubule targeting of FAs. Taken together, these results suggest that Prickle1 promotes FA disassembly during the retraction processes of cell polarization and migration.
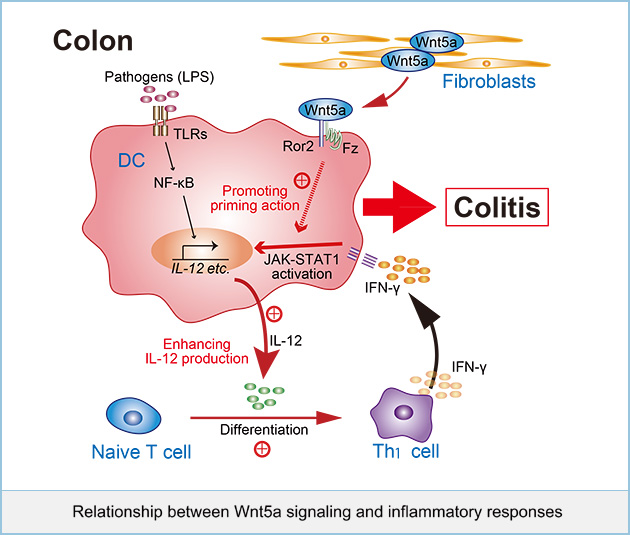
 Relationship between Wnt5a signaling and inflammatory responses
Wnt5a signaling has been recently suggested to be involved in inflammatory diseases. It was shown that expression of Wnt5a and Fz5 can be induced through Toll-like receptors (TLR) stimulated by Mycobacterium tuberculosis (M. tuberculosis) or lipopolysaccharide (LPS) in macrophages, and Wnt5a signaling is involved in producing pro-inflammatory cytokines, such as interleukin (IL)-12 and IL-6. Wnt5a has been detected in granulomatous lesions in the lungs of patients with M. tuberculosis, bone marrow macrophages in septic patients, and macrophages accumulated within the intima of atherosclerotic patients. However, it is still unclear how Wnt5a regulates inflammatory responses physiologically and how its aberrant signaling leads to inflammatory diseases.
We examined the role of Wnt5a signaling in intestinal immunity using conditional knockout mice for Wnt5a and its receptor Ror2. Removing Wnt5a or Ror2 in adult mice did not show clear phenotypes, seems to be healthy macroscopically. However, in these CKO mice, dextran sodium sulfate (DSS)-induced colitis were suppressed. It also attenuated the DSS-dependent increase in inflammatory cytokine gene expression and decreased interferon-γ (IFN-γ)-producing CD4+ Th1 cells in the colon. Wnt5a was highly expressed in stromal fibroblasts in ulcerative lesions in the DSS-treated mice and inflammatory bowel disease patients.
Dendritic cells (DCs) isolated from the colon of Wnt5a and Ror2 deficient mice reduced the ability to differentiate naive CD4+ T cells to IFN-γ-producing CD4+ Th1 cells. In vitro experiments demonstrated that the Wnt5a-Ror2 signaling axis augmented the DCs priming effect of IFN-γ, leading to enhanced lipopolysaccharide (LPS)-induced interleukin (IL)-12 expression. Taken together, these results suggest that Wnt5a promotes IFN-γ signaling, leading to IL-12 expression in DCs, and thereby inducing Th1 differentiation in colitis.
It has been reported that the β-catenin dependent pathway is activated in intestinal DCs and is required for the secretion of immunosuppressive cytokines. In contrast, our results suggested that the β-catenin independent pathway thorough Wnt5a-Ror2 signaling augmented intestinal inflammatory responses in DCs. Therefore, it is intriguing to speculate that in addition to the previously reported action of the β-catenin dependent pathway, which triggers immunosuppressive responses, Wnt signaling may also trigger inflammatory responses through the β-catenin independent pathway in intestinal DCs. Tumorigenesis has been shown to be associated with chronic inflammation, and Wnt5a signaling is also involved in tumor growth and metastasis. Therefore, Wnt5a signaling might be a therapeutic target for human diseases associated with inflammatory disorders, in addition to cancer in future.
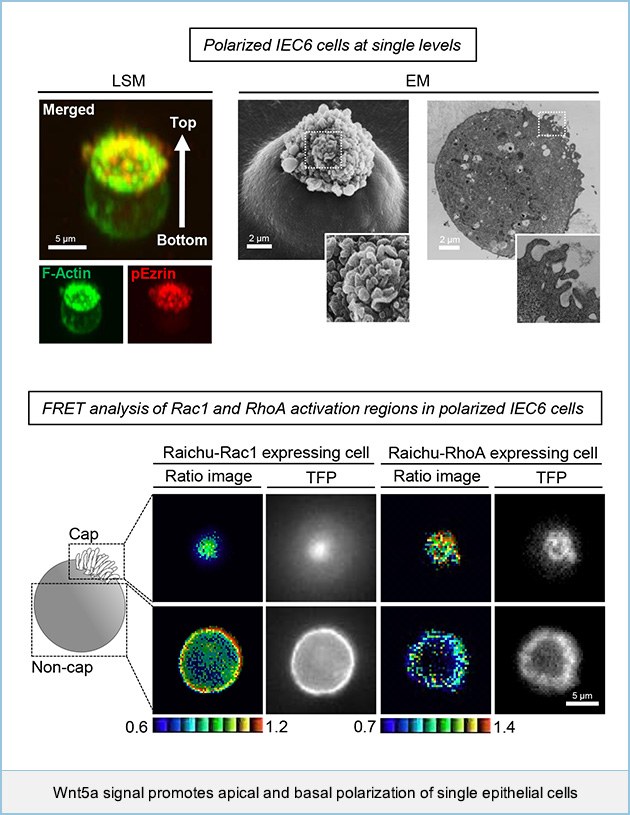
 Wnt5a signal promotes apical and basal polarization of single epithelial cells
Epithelial cells are organized into multicellular sheets of tubules that form distinct apical and basolateral (AB) compartments, which is known as epithelial polarization. It is well known that ECM-mediated signaling is important for the establishment of epithelial polarity. However, physiological cues and molecular mechanisms for ECM-induced AB polarization of normal epithelial cells remain unclear. We show that rat intestinal epithelial cells (IEC6 cells) can polarize at a single cell level in Matrigel- and Wnt5a signaling-dependent manner. When IEC6 cells were seeded on Matrigel, single cells formed an F-actin cap on the upper cell surface, where apical markers were accumulated, and a basolateral marker was localized to the rest of the cell surface region. Rac1 activity in the non-cap region was higher than that in the cap region, whereas Rho activity increased towards the cap region. Wnt5a signaling activated and inhibited Rac1 and RhoA, respectively, independently through Tiam1 and p190RhoGAP-A, which formed a tertiary complex with Dishevelled. These results suggest that Wnt5a promotes the AB polarization of IEC6 cells through the regulation of Rac and Rho activities in a Matrigel adhesion-dependent manner. This study revealed that Wnt5a promotes the AB polarization of IEC6 cells through the spatial regulation of Rac and Rho in an a manner dependent on the adhesion to ECM.
 5. Inside-out and outside-in regulation of Wnt signaling
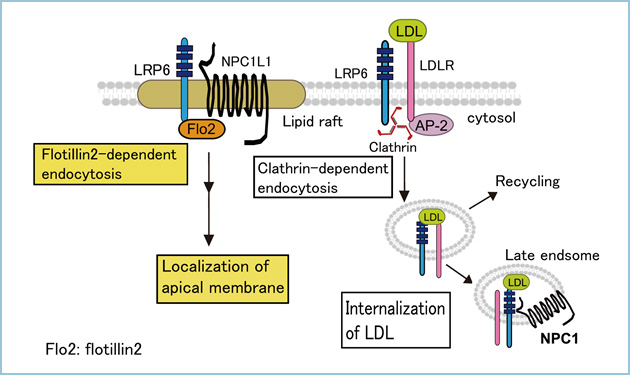
LDL switches the internalization route of LRP6 and determines the destination of NPC1L1
LRP6 was originally identified as a coreceptor of the Wnt signaling pathway and has been shown to be involved in LDL transport. In polarized hepatocytes, many apical proteins are sorted to the basolateral membrane and then internalized and transported to the apical bile canalicular membrane. We examined the distribution and trafficking routes of LRP6 in a polarized hepatocyte model (HepG2 cells). We found that once LRP6 is trafficked to the basolateral membrane, LRP6 is transcytosed with Niemann?Pick type C1-like 1 (NPC1L1) protein to the apical bile canalicular membrane in a flotillin-dependent manner in the absence of LDL. Hepatic NPC1L1 may play a role in the absorption of cholesterol from the bile. In addition, LRP6 was internalized in a clathrin-dependent manner and colocalized with NPC1 in the presence of LDL, which decreased trafficking of NPC1L1 to the apical membrane. These results suggest that LRP6 is internalized via different endocytic routes, depending on ligand availability, and that LRP6 controls the intracellular destination of NPC1L1.
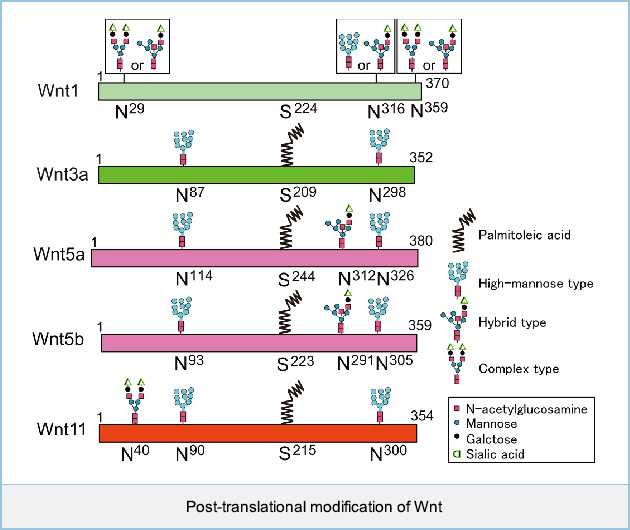
 Post-translational modifications of Wnt1, Wnt3a, Wnt5a, Wnt5b, and Wnt11
Wnts are glycan- and lipid-modified morphogens that are important for cellular responses, but glycan profiles of Wnts are not known. Analysis of the glycosylation pattern of Wnt1 by deglycosylation enzymes revealed that Wnt1 is modified with high-mannose-type or hybrid-type glycan (Asn316) and two complex-type or hybrid-type glycans (Asn29 and Asn 359). Mass-spectrometric analyses revealed that Wnt3a is modified with two high-mannose-type glycans (Asn87 and Asn298), that Wnt5a is modified with two high-mannose-type glycans (Asn114 and Asn326) and hybrid-type glycan (Asn312), that Wnt5a is modified with two high-mannose-type glycans (Asn93 and Asn305) and hybrid-type glycan (Asn291), and that Wnt11 is modified with complex-type glycan (Asn40) and two high-mannose-type glycans (Asn90 and Asn300). Wnt5a, Wnt5b, and Wnt11 are modified with palmitoleic acid at Ser244, Ser223, and Ser215, respectively, which is necessary for the interaction with Wntless and Frizzled receptors, as well as Wnt3a. Wnt1 also has a conserved serine residue (Ser 224) which is the site of palmitoylation for Wnt3a (Ser 209).
 Selective regulation of Wnt signaling by receptor-mediated endocytosis
There are at least two Wnt signaling pathways dependent on or independent of β-catenin. A group of Wnts, including Wnt1 and Wnt3a, activate the β-catenin pathway. In this pathway, the binding of Wnt3a to Frizzled (Fz) and low density lipoprotein receptor-related protein 6 (LRP6) triggers the phosphorylation of LRP6 and induces the recruitment of the Axin complex to phosphorylated LRP6. The phosphorylation of LRP6 occurs in the lipid raft microdomain and that Wnt3a induces the internalization of Fz and LRP6 through caveolin-mediated endocytosis. Caveolin may act on the Axin complex to dissociate β-catenin from Axin, resulting in the accumulation of β-catenin. Another group of Wnts, including Wnt5a and Wnt11, activate the β-catenin-independent pathway, which primarily modulates cell movement, as initially observed during embryogenesis. In this pathway, Wnt5a and Wnt11 binds to receptor tyrosine kinase-like orphan receptor 2 (Ror2) and receptor-like tyrosine kinase (Ryk), respectively, in addition to Fz. They induce the internalization of receptors in a clathrin-dependent manner and activate the small G protein Rac. We are currently analyzing the molecular mechanisms how endocytic processes of receptors are involved in the specific activation of the β-catenin-dependent and -independent pathways.
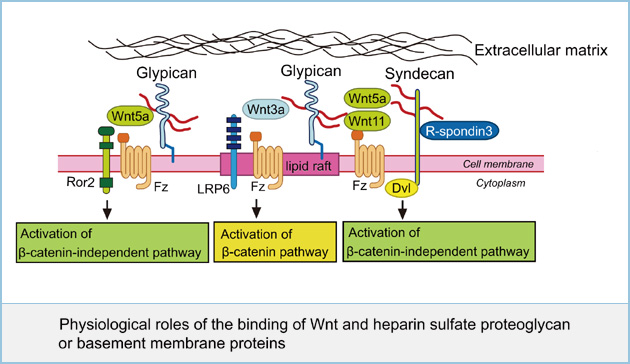
 Physiological roles of the binding of Wnt and heparin sulfate proteoglycan or basement membrane proteins
Heparin sulfate proteoglycans (HSPGs) are localized mainly on the cell surface and in extracellular matrix (ECM). The major cell membrane HSPGs are the transmembrane syndecans (SDCs) and the glycosylphosphatidylinositol (GPI) anchored glypicans (GPCs). Although HSPGs affect the β-catenin-dependent and -independent pathways of Wnt signaling, how they regulate distinct Wnt pathways is not clear. We found that GPC4 is present in both lipid raft and non-lipid raft microdomains and that GPC4 in lipid raft microdominas activates the Wnt3a-dependent β-catenin pathway and GPC4 in non-lipid raft microdomains activates the Wnt5a-dependent β-catenin-independent pathway. In contrast, SDCs appear in non-lipid raft microdomains. It was also reported that SDC4 binds to R-spondin3 and cooperates with Wnt5a and Fz7 to activate JNK in Xenopus embryos through clathrin-mediated endocytosis. These results suggest that GPC4 and SDC4 concentrate Wnt3a and Wnt5a to the vicinity of their specific receptors in different membrane microdomains, which regulates distinct Wnt signaling. To further understand the mechanism of the efficient binding of Wnts to their receptors, we are investigating basement membrane proteins and HSPGs which interact with Wnts and regulate Wnt signaling pathway.
 Regulation of polarized sorting of Wnts and their receptors in MDCK cells
The mechanism underlying how Wnts and their receptors are sorted to their specific destinations is largely unknown. We examined the polarized sorting of Wnts and their receptors. The apical and basolateral secretion of Wnts are regulated by different mechanisms. The basolateral secretion of Wnt3a and Wnt5a required clathrin and adaptor protein 1 (AP-1) in polarized epithelial cells. On the other hand, glycosylation processing at Asn40 and galectin-3 are required for the apical secretion of Wnt11. Wntless (Wls) is essential for Wnt secretion, and is trafficked to the basolateral membranes by clathrin and AP-1 from the trans-Golgi network. Frizzled2 mainly localized to the basolateral membranes in MDCK cells. LRP6 and Ror2, which are receptors for Wnt3a and Wnt5a, respectively, were also trafficked to the basolateral membranes using clathrin and AP-1, but their sorting did not require Wls. The signaling pathways of Wnt3a and Wnt5a were strongly activated when both Wnts stimulate the basolateral side. These results indicate that the appropriate Wnt receptors are sorted to the same polarized fractions as the corresponding Wnts, thereby leading to efficient activation of their signals.
 
|
|