

INDEX
- 1. The principle of voltage sensor
- 2. Molecular mechanisms and structural biology of VSP
- 3. Physiological roles of voltage-gated proton channels in blood cells and microglia in brain
- 4. Molecular tool development for the visualization of membrane potential and manipulation of phosphoinositides.
- 5. Elucidating new roles of membrane potential signals in biological phenomena
- 6. Turnover of voltage-gated ion channels in mammalian brain related to pathophysiology and ageing.
- 7. Fundamental studies of visual function recovery based on artificial retina. (Dr. Tomomitsu Miyoshi)
-
1. The principle of voltage sensor
For a long time, studies on the mechanisms of neuron excitation and impulse transmission have been a major theme in physiology. In the mid-20th century, Huxley and Hodgkin discovered that sodium ions play an essential role in the formation of action potentials in the giant axon of the squid. They quantitatively measured the ion currents and demonstrated the following:
(1) Na+ and K+ permeate the plasma membrane through different pathways.
(2) The opening and closing of these pathways are quantitatively described by the function of voltage-gated ion channels, which are dependent on time and membrane potential. Action potentials were numerically reconstituted based on these elements.Ion channels, including voltage-gated sodium channels and potassium channels, were characterized in detail using toxins and by measuring microscopic currents by patch-clamp technique. In the late 20th century, molecular cloning techniques led to the identification of these ion channels. In the 21st century, the structure of voltage-gated ion channels at the atomic level was resolved by X-ray crystallography, leading to a clearer understanding of the channels.
In the years 2005 and 2006, we discovered two unique voltage-dependent membrane proteins (Murata et al, Nature, 2005; Sasaki et al, Science,2006). One of the proteins, identified from ascidians (Ciona intestinalis), was named Ci-voltage sensing phosphatase (Ci-VSP). This protein consists of two modules: a voltage sensor and a phosphatase. Upon depolarization the voltage sensor undergoes a conformational change, which is coupled to the enzymatic activity of the phosphatase. This enzyme dephosphorylates phosphatidylinositol, which are involved in various signalling functions within the cell. Therefore, we found that VSPs are responsible for the conversion of electrical signals to chemical signals.
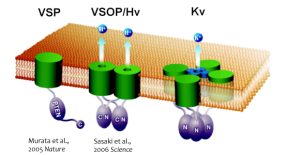
Fig.1 The new types of voltage sensor proteins
VSP: Voltage-sensing phosphatase,VSOP: Voltage sensor domain only protein (also known as Hv1, Voltage-gated proton channel),Kv: Voltage-gated potassium ion channel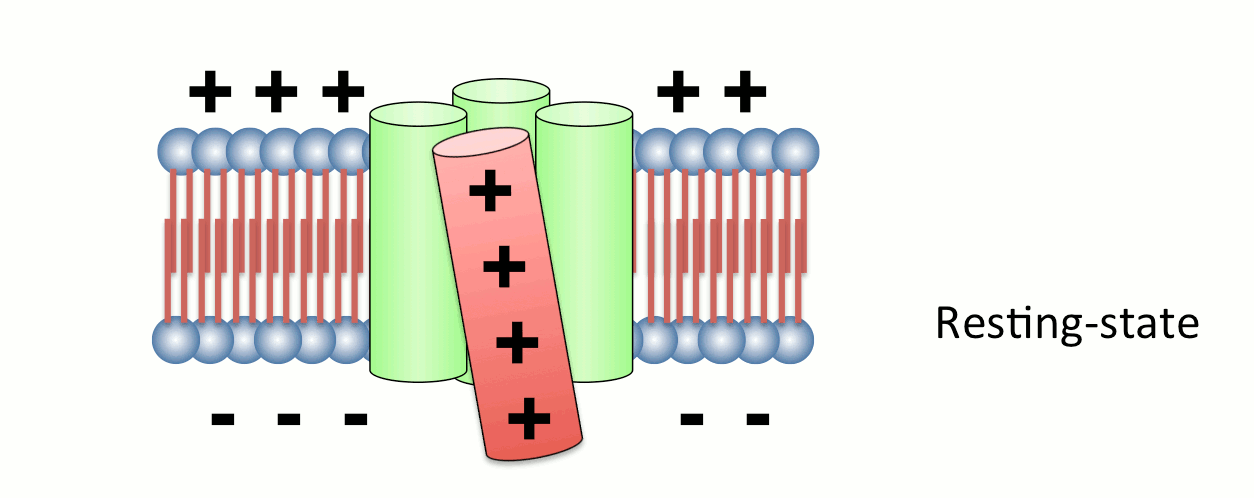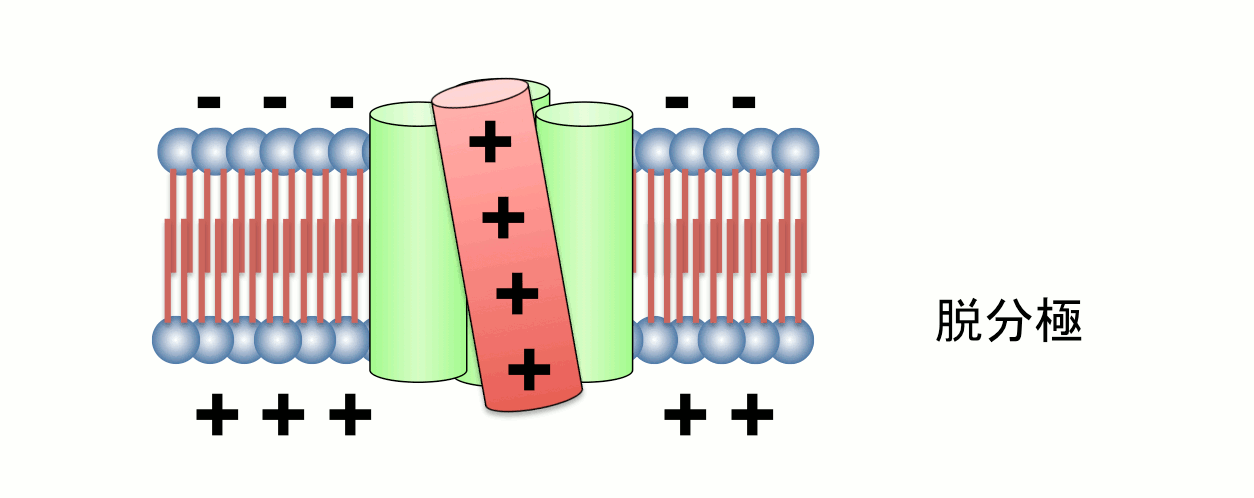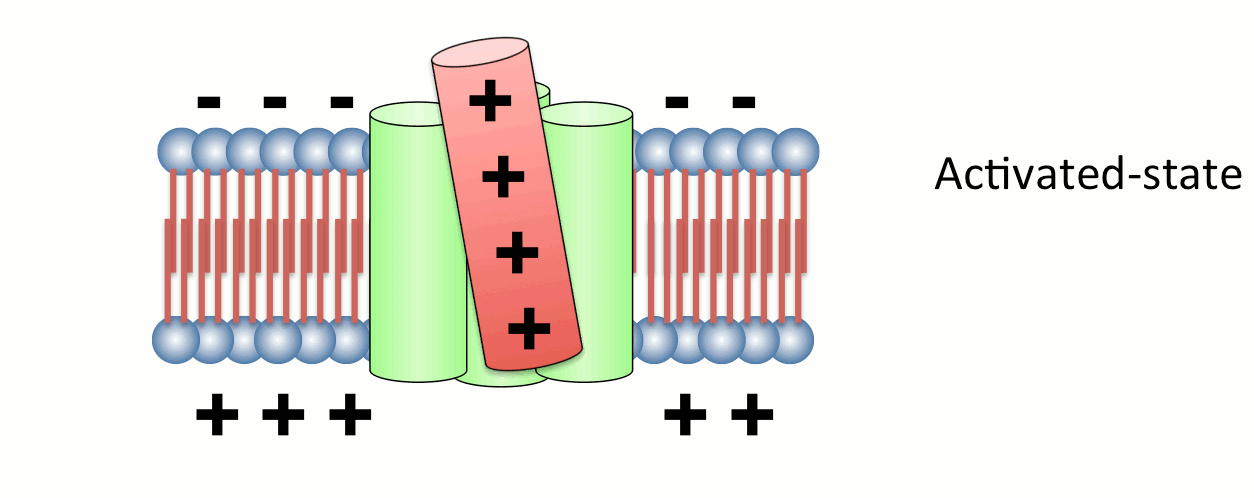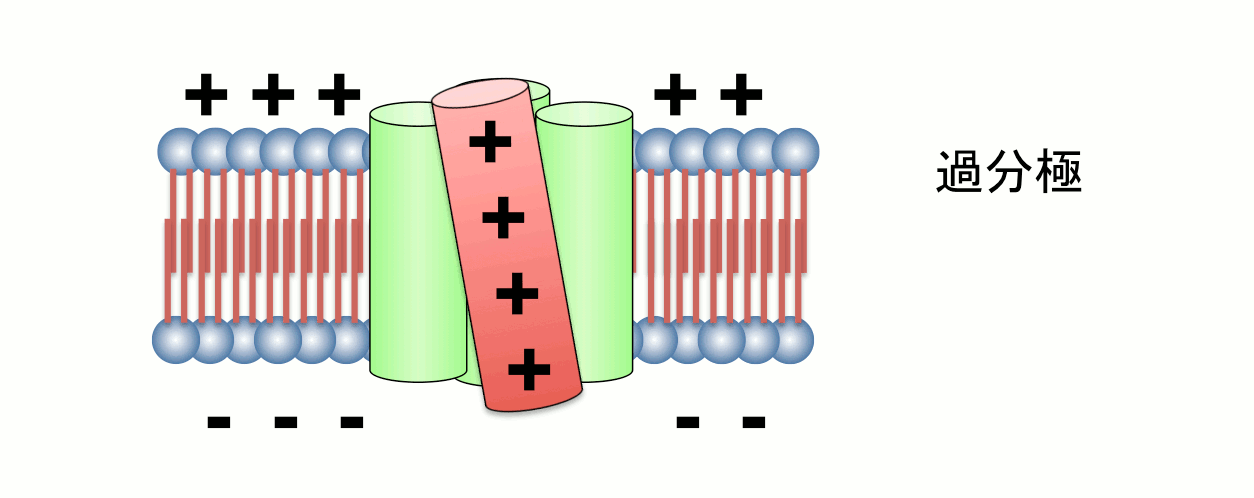
Fig.2 Operation of voltage sensor domain.
Among four transmembrane helices, the fourth segment, called S4, has positive charges to sense transmembrane voltage gradient and trigger motion of the whole protein structure in response to voltage change. Animation: by the courtesy of Dr. Kohei Takeshita, RIKEN.The other protein, named VSOP/Hv1 (VSOP=voltage sensor domain only protein), as the name indicates, consists only the voltage sensor domain. In addition, this protein lacks an authentic pore domain but exhibits proton-selective and voltage-dependent conductance. The voltage sensor domain of VSOP/Hv1 has roles in both the voltage-dependent gating and proton permeation. Both "gating" and "ion permation" are two central questions in the field of ion channels. Thus, in this sense, VSOP/Hv1 provides a good model to study the general principle of ion channel.
The Hv channel consists of two monomers that associate through cytoplasmic coil-coil interactions (Koch et al, PNAS, 2008, in collaboration with Dr. Peter Larsson). X-ray crystallographic analysis of the cytoplasmic region of VSOP/Hv1 revealed that the mechanism of opening and closing is precisely controlled by the coil-coil structure interactions, which occur at normal body temperature (Fujiwara Y. et al, Nat. Communi., 2011).Additionally, VSOP/Hv1 is sensitive to extracellular metal cations, and in particular, zinc ions, and the critical amino acids for zinc sensitivities have been identified. This zinc sensitivity is crucial for fertilization of human sperm which expresses VSOP/Hv1. Before ejaculation, seminal fluid surrounding human sperm contains high concentration zinc which persistently inhibits VSOP/Hv1 activity of sperm. But, after ejaculation of sperm into female reproductive space, sperm’s environmental zinc concentration is diluted resulting in de-repression of VSOP/Hv1 activity leading to activation of sperm due to cytoplasmic alkalization.
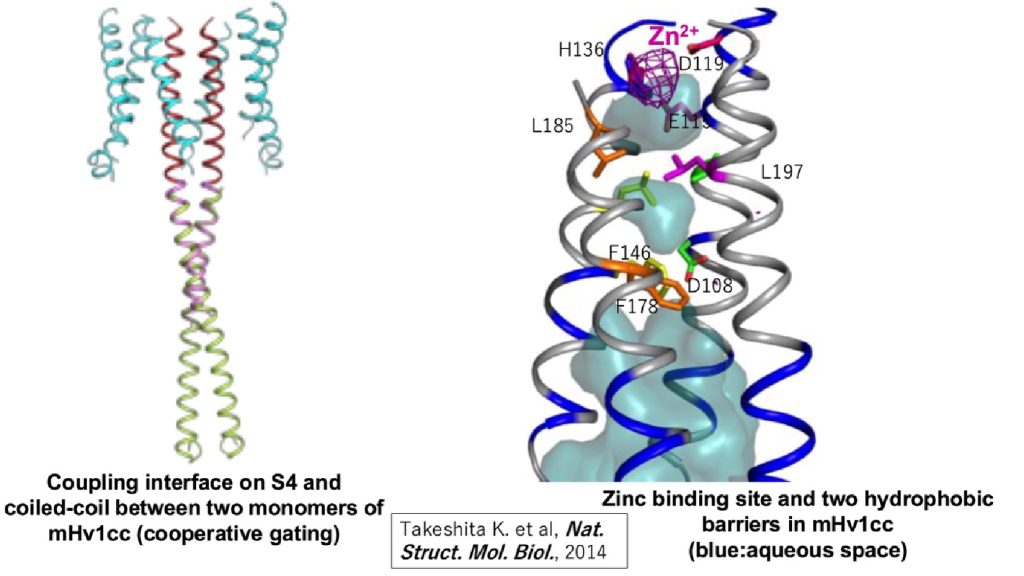
Fig.3 The structure voltage-gated proton channel (mHv1cc: mouse VSOP/Hv1-CiVSP chimera)
- 1. How is the phosphatase activity of VSP controlled by the function of the voltage sensor?
- 2. How do changes in the membrane potential make VSOP/Hv1 conductive to protons?
To address these questions, we integrate techniques from electrophysiology, fluorometry, genetic code expansion, and molecular dynamics simulation. This comprehensive approach allows us to explore the details of the molecular mechanisms underlying structure-function relationships of these proteins.
-
2. Molecular mechanisms and structural biology of VSP
To date, many studies highlight the roles of tyrosine phosphatases and PTEN in the development of cancer and metabolic disorders. Interestingly, the cytoplasmic enzyme region of the VSP share structural similarities with these enzymes but possesses the unique feature of having its enzymatic activity being controlled by changes in membrane potential. Understanding the mechanisms underlying this regulation could provide valuable insights into how tyrosine phosphatases and PTEN contribute to the development of cancer and its related diseases.
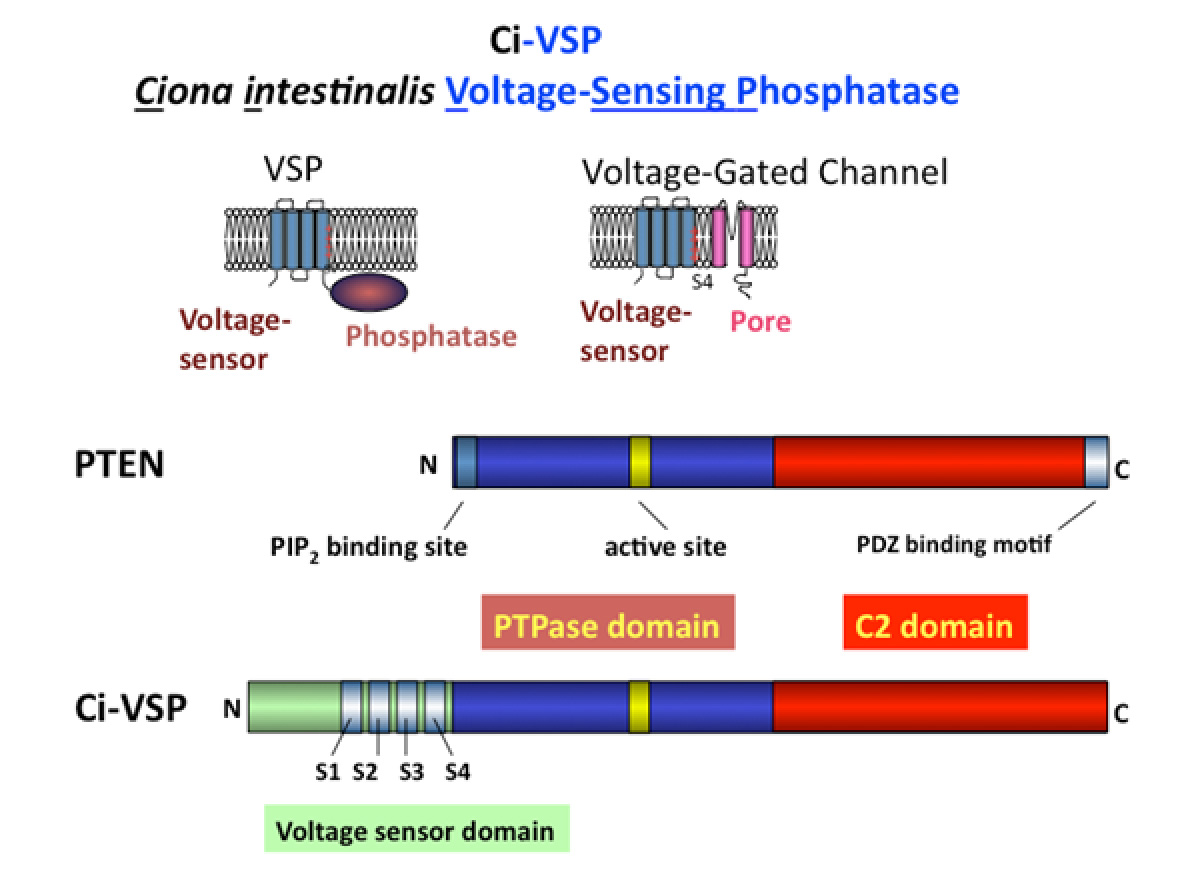
Fig.4 VSP and PTEN The enzymatic domain of VSP shares significant structural similarity with PTEN, both enzymes targeting phosphoinositides as substrates (Iwasaki H et al.,PNAS, 2008; Matsuda M et al, JBC, 2011). A unique feature of VSP is its ability to alter substrate specificity in response to changes in membrane potential, a feature not a common among most enzymes (Kurokawa T et al, PNAS, 2012). Our extensive studies on the molecular mechanisms underlying the coupling between the voltage sensor and the enzyme have identified key regions involved in this interaction. One critical site is the hydrophobic spine, a site where the enzyme associates with the membrane (which also turned out to be conserved among many phosphoinositide phosphatases including PTEN and MTMR2)(Kawanabe A. et al, eLife, 2018). The second site is situated at the lower region of the S4 segment, which contains positive charges important in sensing the transmembrane potential. We found that these two sites interact upon membrane depolarization (Mizutani N. et al, PNAS,2022).Notably, the lower part of the S4 segment shares structural similarity with the N-terminal end of PTEN, known to be critical for the regulation of enzymatic activity. This suggests that the voltage sensor of VSP “hijacks” the innate regulatory mechanism of enzymatic activity in PTEN, which lacks a voltage sensor domain.
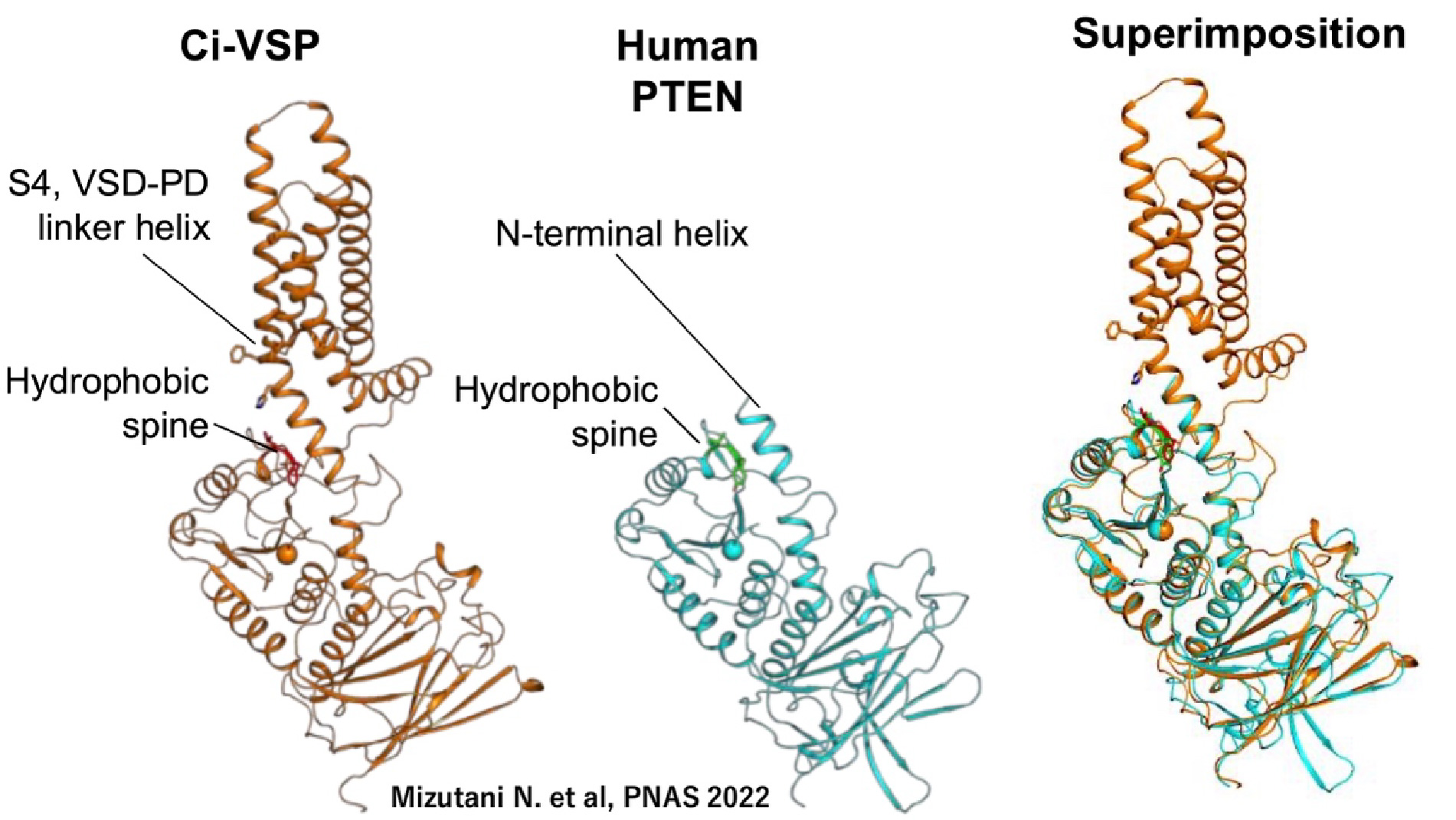
Fig.5 Key interdomain interactions for coupling in VSP (Mizutani N. et al, PNAS, 2022)
-
3. Physiological roles of voltage-gated proton channels in blood cells and microglia in brain
VSOP/Hv1(Sasaki M. et al, Science, 2006) is conserved from marine invertebrates to human. In mammals these channels are expressed in blood cells such as macrophages, neutrophils, and microglia. In many mother cells, including osteoclasts, ATP hydrolysis by proton pump (V-ATPase) results in the production of protons, which are then transported to the extracellular side. It is well known that Hv channels are able to export large quantities of protons from the cell more accurately than proton-pump pathways. In most cases, Hv channels are only activated when the membrane potential is more positive than the reversal potential for proton flux. This is due to intrinsic properties of pH-regulated gating of Hv channels. Vectoral transport of proton to extracellular side can be achieved without requiring the use of ATP.
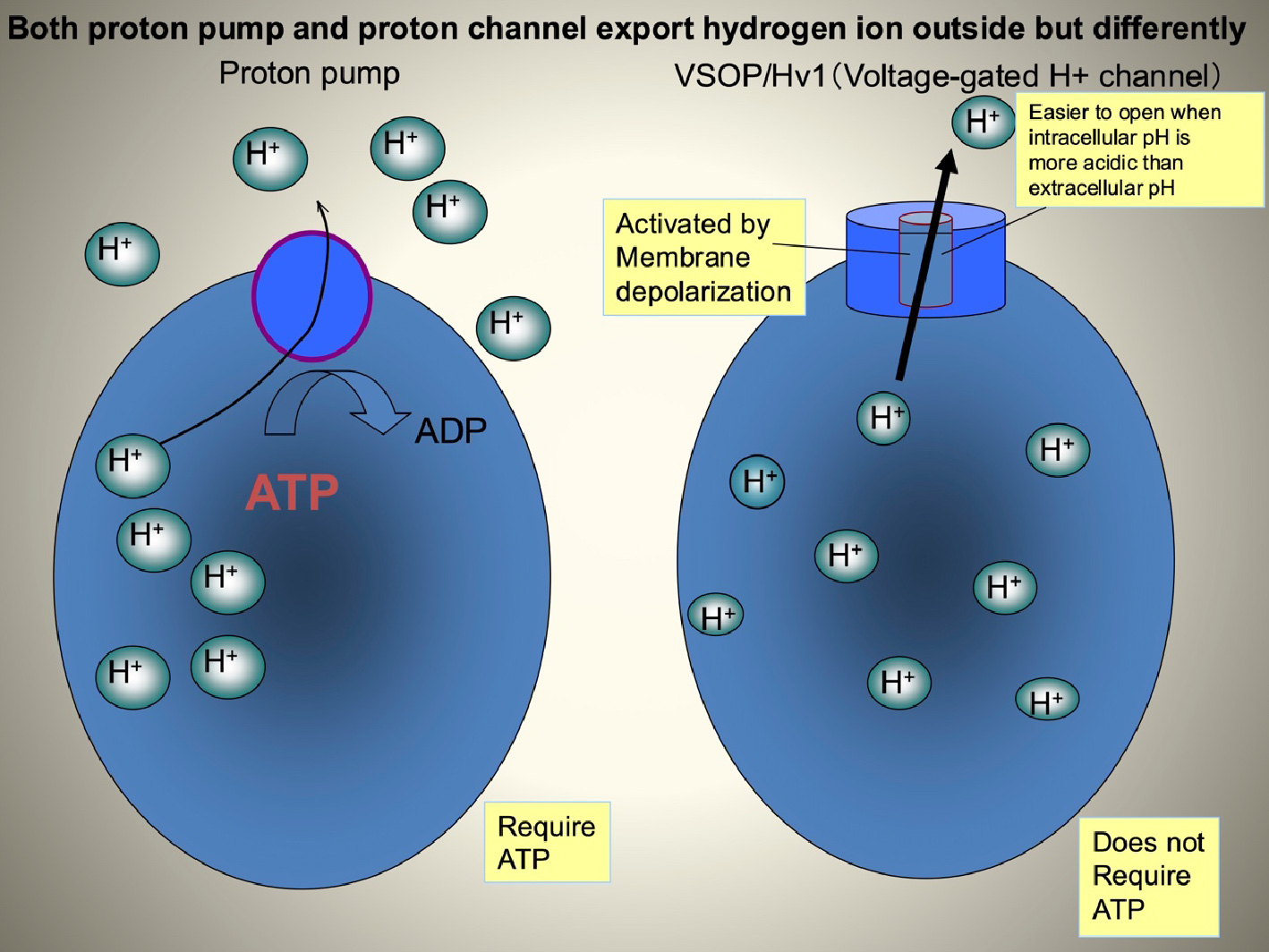
Fig.6 The difference between voltage-gated proton channel and proton pump Electrophysiological studies have suggested that Hv channels play a role in promoting the production of reactive oxygen species (ROS) in phagocytic cells. Initially, superoxide anions (O2-) are generated by NADPH oxidase, a protein complex comprised of the membrane protein gp91 and other auxiliary subunits. The generated O2- are then secreted into the lumen of phagosomes or to the extracellular side. However, continued accumulation of O2- can lead to an increase in cell membrane potential due to charge imbalance, which subsequently inhibits NADPH oxidase activity and reduces O2- production. Hv channels help correct this charge imbalance by transporting protons, thereby maintaining continued ROS production.
In our analysis of VSOP/Hv knockout mice, we found that the Hv channel controls both cytoplasmic pH and membrane potential, by which is through the proton transport, sustaining ROS production (Okochi Y. et al, BBRC, 2009, El Chemaly et al, JEM, 2010). This regulation has been linked to various unexpected phenotypes, including alterations in neutrophil chemotaxis, glucose metabolism, and behavior (Okochi Y, et al, J. Leuk. Biol., 2020; Kawai T., et al, FASEB J. 2020 in collaboration with Dr. S. Kihara’s lab; Kawai T., et al J. Neurochem, 2021 in collaboration with Dr. K. Sakimura, Dr. M. Abe and Dr. T. Miyakawa). Notably, we found that some phenotypic changes in the VSOP/Hv1-deficient mice are closely associated with ageing, and so we are now collaborating with Dr. Oginuma, a fish geneticist at RIKEN, to explore the relationship between VSOP/Hv1 function and longevity. We have also been working on marine invertebrates and unicellular organisms which utilize VSOP/Hv1 for cellular homeostasis in collaboration with researchers of earth science and marine science (based on the grant, “pH Biology”).
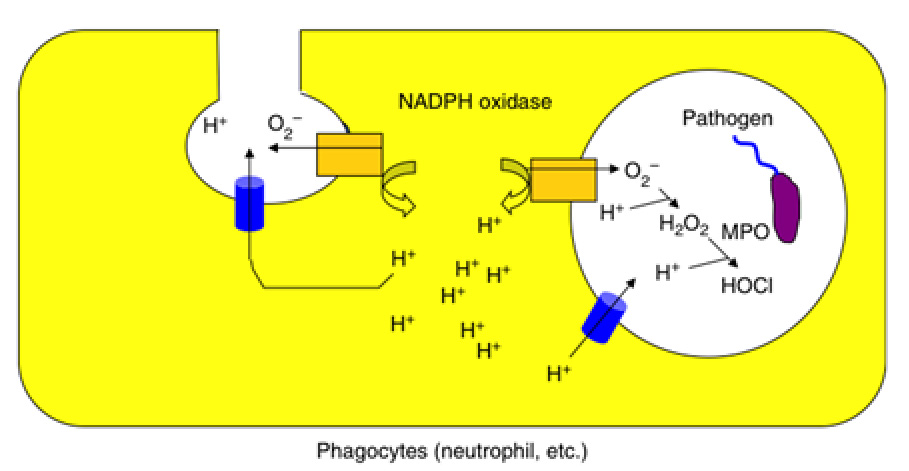
Fig.7 The physiological role of voltage-gated proton channel on phagocyte such as neutrophil and macrophage -
4. Molecular tool development for the visualization of membrane potential and manipulation of phosphoinositides.
VSP is a valuable resource for developing molecular tools in neuroscience and cell biology. Understanding neuronal circuits, which are fundamental to brain function, requires precise measurements of individual neuron activities. Traditional electrophysiological approaches, such as using glass electrodes to cells and tissue cultures, are relatively tedious. Additionally, it will be highly useful to visualize the membrane potential and behaviour of multiple neurons using non-invasive methods.
Historically, potential-sensitive organic dyes have been used as voltage probe, but these dye-based probes are challenging to target specific cells and often produce weak signals. Genetically encoded voltage indicators (GEVI) have been developed by linking the voltage sensor of VSP to fluorescent proteins, providing a more targeted and efficient solution. One notable example is the Mermaid (a combination of proteins derived from marine organisms), including Mermaid2 which was developed by Dr. H. Tsutsui (currently an associate professor in JAIST). Significant changes in fluorescence were measured in the imaging of cultured cells, such as single neuron and cardiac cells (Tsutsui T et al., Nature Methods, 2008) and later cerebral cortex of in vivo mouse (Tsutsui T. et al., J. Physiol. 2013 PMID: 23836686).
Approximately half of GEVIs worldwide is based on VSP. The rest of the GEVIs are mainly rhodopsin-based. VSP-based GEVIs are particularly advantageous for in vivo live imaging using multi-photon fluorescence microscopy. Therefore, VSP-based GEVIs offer an alternative method of non-invasive approaches for studying neuronal circuitry and other cellular activities, to enhance our understanding of complex biological processes.
VSP also serves as a valuable tool for manipulating plasma membrane PI(4,5)P2. Please see a couple of papers for examples of VSP as a molecular tool in studies of phosphoinositide biology (Tsutsui H, Mizutani N, Okamura Y (2021) “Engineering voltage sensing phosphatase (VSP)”. Methods in Enzymology ISSN:0076-6879, 654:85-114; Okamura Y, Yoshioka D. “What voltage-sensing phosphatases can reveal about the mechanisms of ion channel regulation by phosphoinositides”. Biochem Soc Trans. 2023 Apr 26;51(2):827-839. PMID: 37052219.). Furthermore, using both wild-type and engineered VSPs, we have recently discovered previously unappreciated roles of PI(4,5)P2 in transporter protein (Mizutani N., Physiol. Rep. 2019) and ligand-gated ion channels (Okamura Y., 2024 Gordon Research Conference, keynote lecture).
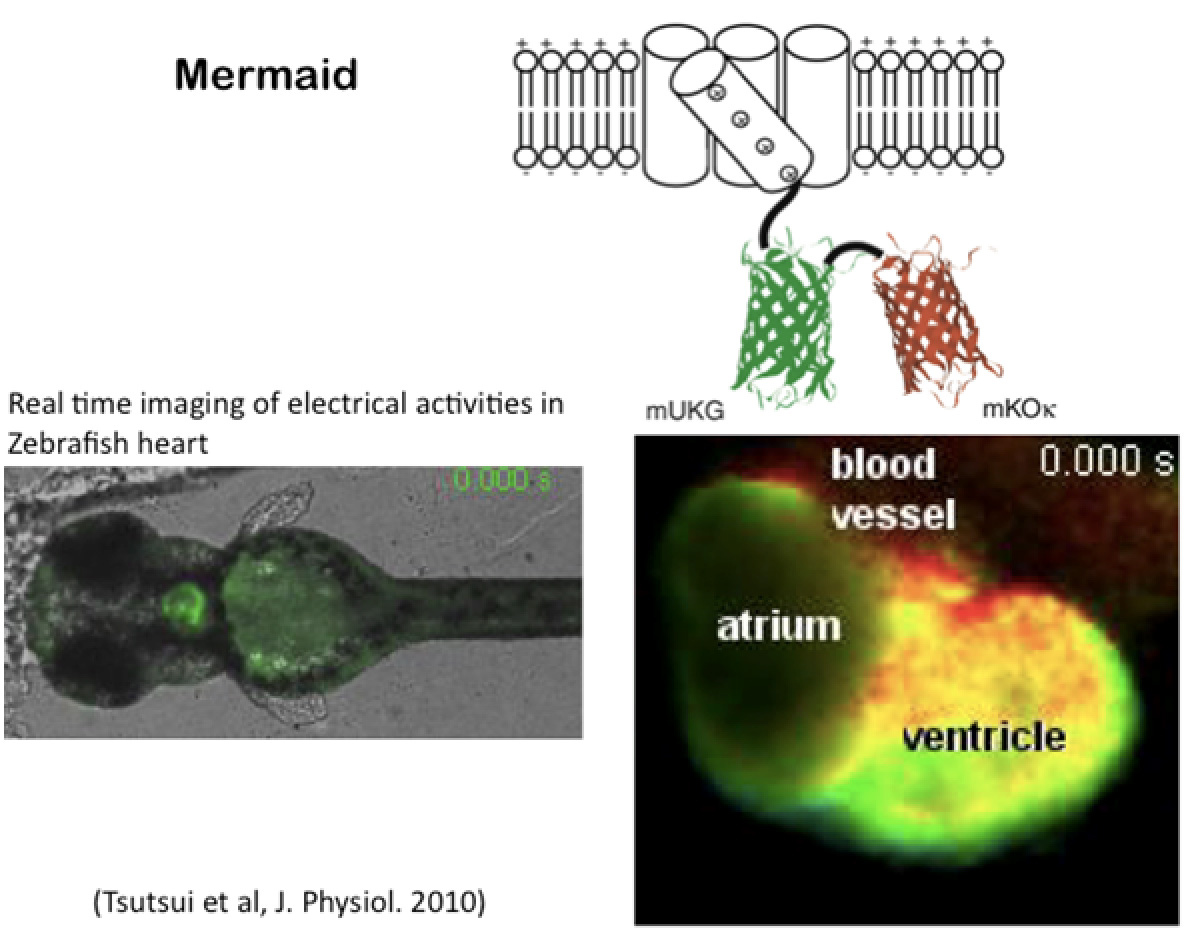
Fig.8 Example for visualizing membrane potential based on VSP. Right lower panel shows snapshot of FRET-based imaging movie of in vivo heart at the whole animal of zebrafish. -
5. Elucidating new roles of membrane potential signals in biological phenomena
Research on voltage sensor proteins has led to the discovery of new molecular pathways linked to changes in membrane potential, prompting a reassessment of physiological roles of membrane potential. Notable examples include the membrane potential’s role in the polyspermy block during fertilization, involvement in the left-right nodal determination, and its function in regeneration processes in non-mammalian vertebrates. Although the precise molecular mechanisms behind these processes remain unclear, they highlight the broad physiological implications of membrane potential changes. Together with our collaborators, we are actively investigating the physiological roles of VSP and other voltage-sensitive proteins to better understand these complex processes.

Fig.9 VSP function during maturation of mouse sperm
(Kawai T. et al. Nature communi. PMID: 39181879); in collaboration with Drs. K. Watanabe, T. Sasaki, J. Sasaki, K. Sakimura, M. Abe, M. Ikawa, H. Miyata). Analysis of knock-in mice with single amino acid mutation in the voltage sensor domain showed that VSP senses membrane potential in immature spermatocytes to establish the phosphoinositide conditions which are required for sperm functions.In VSP, the enzymatic activity changes in response to changing membrane potentials. VSP is expressed in testis, where it plays an important role in sperm maturation. Figure 9 illustrates the physiological role of VSP based on its molecular properties. During sperm maturation, VSP senses the resting membrane potential and provides graded distribution of PI(4,5)P2 across the whole sperm flagellum dependent on the enzymatic activity. This spatial distribution of PI(4,5)P2 is critical for the modulation of PI(4,5)P2-sensitivive sperm-specific potassium channel, Slo3, which in turn regulates the calcium influx through CatSper channel—the main pathway for calcium influx in sperm (Kawai T., et al, PNAS, 2019). Our recent study on knockin mice with alteration of voltage sensor structure showed that nature of voltage sensing of VSP is critical for maturation of sperm function through regulating profiles of phosphoinositide species. These verified that VSP senses membrane potential during maturation of sperm. This study also showed, for the first time, that mammalian VSP ortholog functions as the voltage-sensitive PI(4,5)P2 phosphatase in physiological context (Kawai T., et al, Nature Communi, 2024 PMID: 39181879).
-
6. Turnover of voltage-gated ion channels in mammalian brain related to pathophysiology and ageing.
Neurons are unique in two key aspects: their highly polarized morphology and their longevity, which often match the lifespan of the organism. Understanding how neuronal functions are maintained throughout the whole life is an important biological question, closely related to pathological conditions that involve conditions like demyelination, or psychiatric disorders and ageing. Our research focuses on the ion channel turnover in two critical regions of axons: the axon initial segment (AIS) and the nodes of Ranvier.
The AIS is the site of action potential initiation, a concept first proposed in a seminal study on action potentials that analyzed three subcellular compartments of mammalian neurons (J. S. Coombs, D. R. Curtis and j. C. Eccles (1957) The generation of impulses in motoneurones. J. Physiol. I39:232-249). The nodes of Ranvier (referred to as Nodes from this point on) is the structure unique to myelinated axons, situated between two myelin-covered internodes. Action potentials are regenerated or boosted at the Nodes, effectively ‘hopping’ over electrically insulated internodes—a mechanism known as saltatory conduction, discovered by I. Tasaski and later confirmed by R. Stampfli and A. Huxley.
The AIS and the Nodes share similar morphological and chemical architectures, including clusters of voltage-gated sodium channels and potassium channels, as well as cytoskeletal components. Recent studies have identified many proteins critical for organization of these two structures in myelinated axons, with one of the most critical organizers being the multi-modular adaptor protein, Giant ankyrin G protein (Ank3). Among several isoforms encoded by three genes in mammals (AnkyrinG, R, and B), variants of Ank3 gene have been associated with bipolar disorder, schizophrenia, and autism.
The AIS is also plastic as it can change its length or position in response to synaptic inputs in neurons, which is a mechanism underlying the homeostatic regulation of intrinsic neuronal excitabilities (Kuba H., et al, Nature PMID;20543825 2010, etc). While the Nodes have long been considered as more static structures, recent studies suggest that node lengths can also change in an activity-dependent manner, contributing to neuronal plasticity.
We are using taking two approaches with different spatial and temporal resolutions to understand how voltage-gated ion channels in the AIS and Nodes are regulated throughout the lifespan of neurons. The first approach involves examining the three-dimensional, nanometer scale dynamics of voltage-gated potassium channel, KCNQ2/3 (also known as the classical M-channel, historically studied in the neuromodulation of frog sympathetic neurons) through single molecule imaging.
The second strategy is to visualize ion channels in vivo using fluorescent proteins in a genetically engineered mouse model, which allows us to distinguish between old and newly synthesized proteins. By combining these two approaches, we aim to understand how neuronal excitability is maintained over a lifetime and how disruptions in the homeostatic mechanisms lead to abnormal conditions, including psychiatric disorders.
This project is also related to one fundamental question in biology and evolution: how did the vertebrate-type neuron evolve during chordate evolution? Here, we hope to shed light on this important question by building on from historical studies on urochordate neurobiology and ion channels (Okamura Y., et al, Neuron, 1994).
-
7. Fundamental studies of visual function recovery based on artificial retina. (Dr. Tomomitsu Miyoshi)
Collaborating with ophthalmologists, engineering scientists, and an eyecare company, we have developed artificial retinas (retinal prostheses) to treat degenerative diseases of the photoreceptor cells, like retinitis pigmentosa. We proposed the new retinal stimulation method, Suprachoroidal Transretinal Stimulation (STS), which is safer than the existing methods. We have assessed the function of STS-type prostheses with retinal and brain activities such as intrinsic signals, local field responses, and single-unit activities. Newly designed devices and novel stimulating methods have also been developed.
![]()