

INDEX
-
1. 電位センサーの動作原理
神経が興奮を伝達する仕組みは、長い間、生理学の最重要テーマのひとつとして研究されてきました。20世紀半ばに、ホジキンとハックスレーによりヤリイカの神経でナトリウムイオンが活動電位の形成に本質的な役割を担うことが発見され(Na説)、イオン電流の定量的な計測によって、(1)NaイオンとKイオンが別々の通路を通ること、(2)その通路の開閉は、膜電位と時間に依存して変化することが明らかにされ、電位依存性イオンチャネルの概念が確立しました。その後、様々な研究者により、トキシンへの結合を利用したイオンチャネルの単離や、パッチクランプ法による単一イオンチャネルを流れるpAレベルの微小電流の計測、分子クローニングによって電位依存性イオンチャネルの分子実体が明らかにされました。21世紀に入って、X線結晶構造解析によって、電位依存性カリウムチャネルなどの構造が原子レベルで明らかになるに到って、現在も最先端の研究が続いています。
私たちは、2005年と2006年に、世界に先駆けて、電位依存性イオンチャネルよりもシンプルな構造を有する、2つのユニークな膜蛋白質分子を発見しました(Murata et al, Nature, 2005; Sasaki et al, Science, 2006)。VSP(Voltage-Sensing Phosphatase)は、単一の電位センサーと、単一の酵素が分子内で連結した構造を有し、細胞膜の電位が脱分極側へ上がると電位センサーの構造が変化し、その情報が細胞内の酵素領域に伝わり、ホスファターゼ(脱リン酸化酵素)の活性が出現します。つまり、電気シグナルが化学シグナルに変換されることになります。このホスファターゼ活性は、イノシトールリン脂質(リン脂質の一種で細胞内の様々な信号の制御に関わる)を脱リン酸化します。イノシトールリン脂質は、イノシトール環と呼ばれる糖の構造に共有結合しているリン酸の位置と数によって、そのシグナル分子としての役割が変わるため、この脱リン酸化によって細胞内の情報伝達が変化すると考えられます。
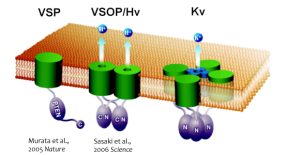
図1. 新規電位センサー蛋白質の発見
VSP : 電位依存性脱リン酸化酵素, VSOP/Hv1 : 電位依存性プロトンチャネル,
Kv : 電位依存性カリウムチャネル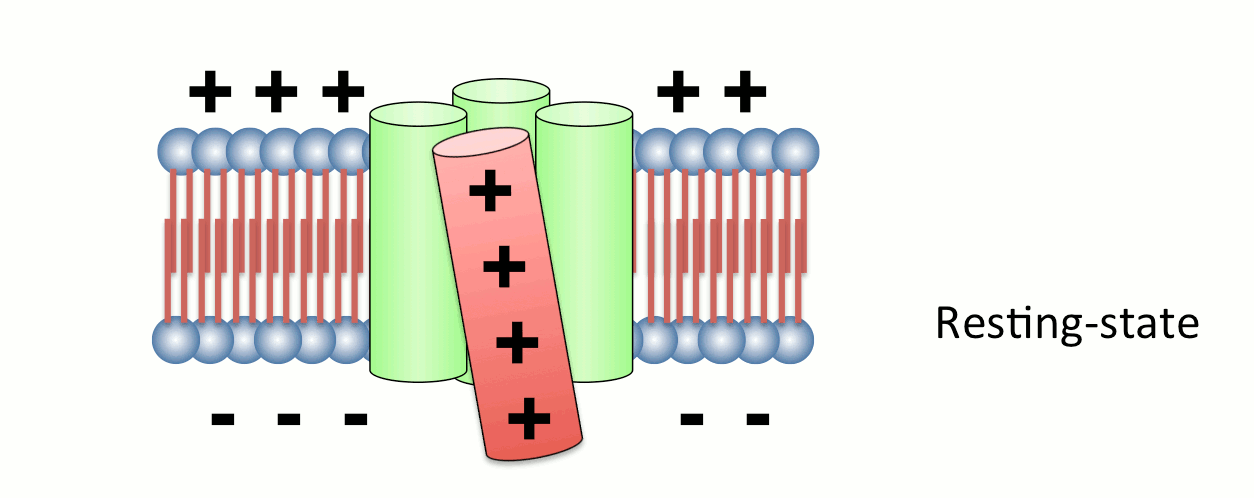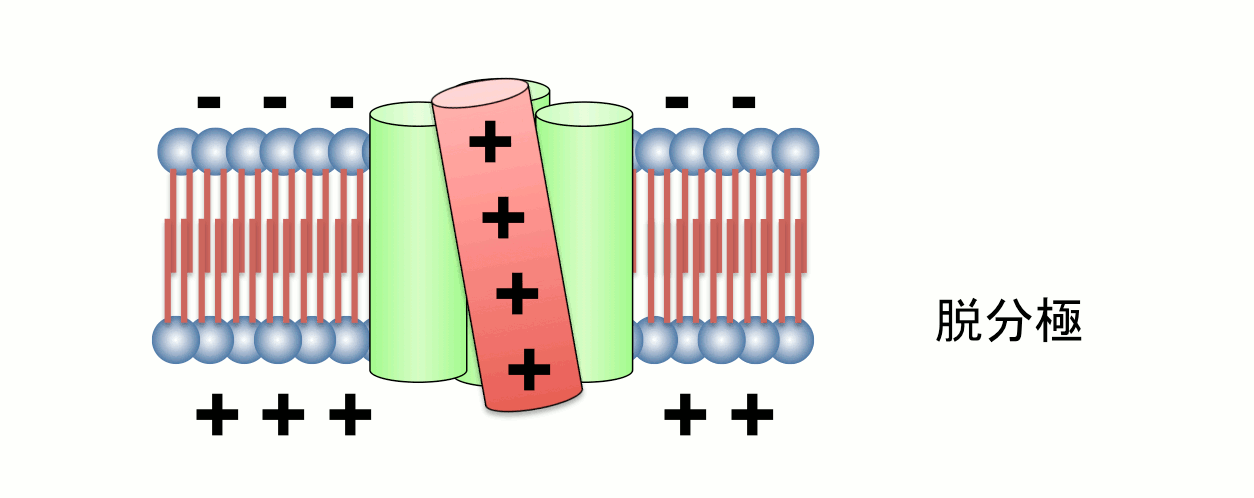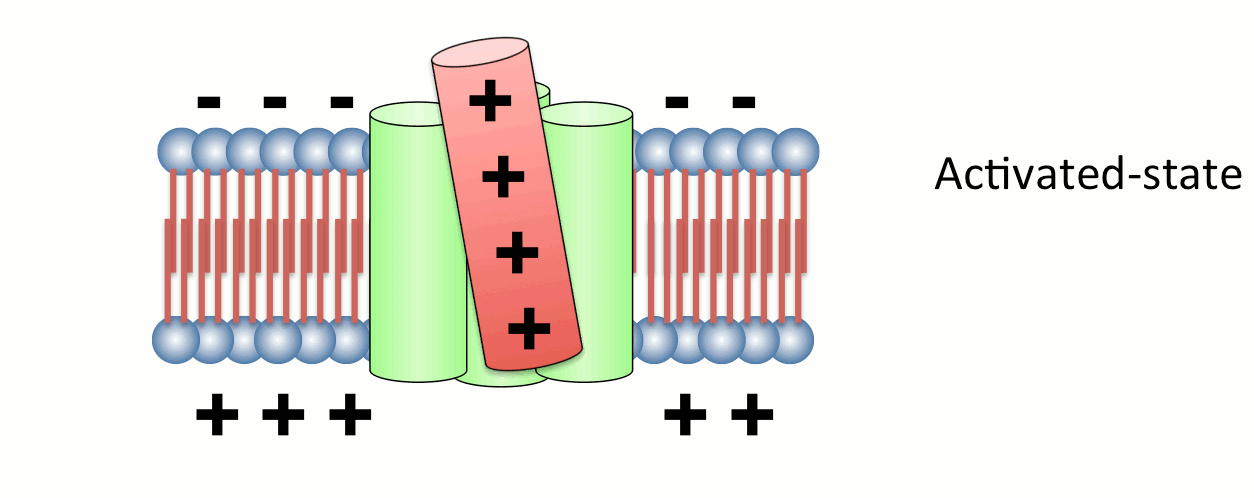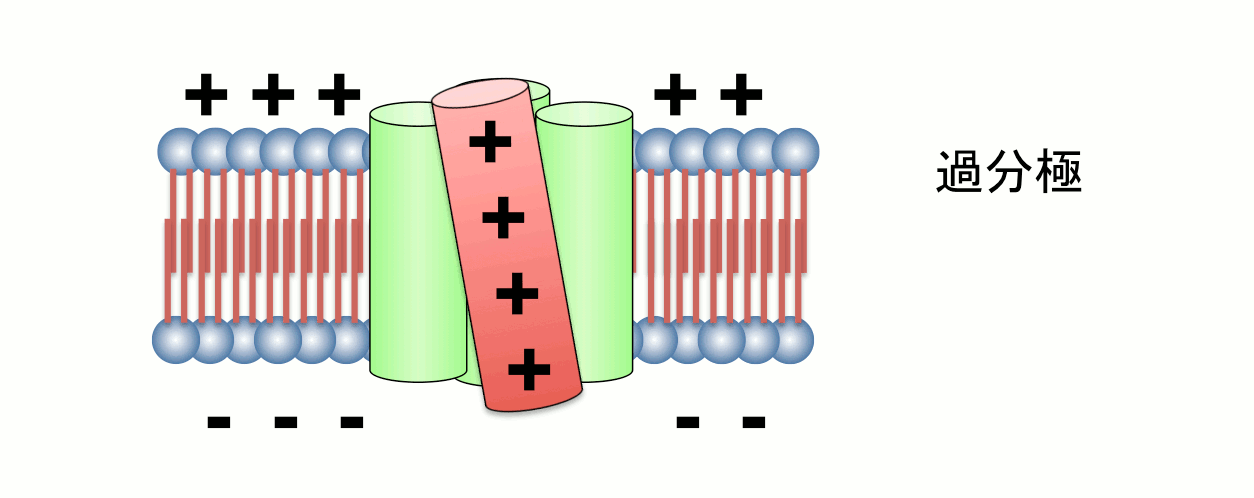
図2. 電位センサーの動き
4つの膜貫通ヘリックスのうち、4番目のセグメント(S4)には膜内外の電位差を感知するプラスチャージを有するアミノ酸が存在する。S4は膜電位変化に応答してタンパク質全体の構造変化を誘導する。
図:理化学研究所・竹下博士の好意による。一方、電位センサードメインのみから成るVSOP(Voltage sensor domain only protein)は、通常のイオンチャネルに見られるイオン透過ドメインをもっていないにも関わらず、水素イオンを通す働きのあるイオンチャネル(電位依存性プロトンチャネル:Hvチャネル)として機能します。膜電位を感知する機能しか持たないはずの電位センサードメインが、VSOPにおいては、膜電位感知と、プロトン透過の、一人二役の性質をもっています。この分子は、細胞内の領域を短く削っても(アミノ酸の長さにして100程度)イオンチャネル分子として機能できることから、哺乳類のゲノムでコードされているすべてのイオンチャネルのうち、最小で機能するイオンチャネルと言えます。
VSOP/Hv1は細胞質内領域にあるコイルドコイルドメインを介して2量体を形成します(Koch et al, PNAS, 2008, Dr. Peter Larssonとの共同研究)。X線結晶構造解析の結果、プロトン開閉機構はコイルドコイルドメインの相互作用によって、しかも体温付近で、精密に制御されていることが分かりました (Fujiwara Y. et al, Nat. Communi., 2011)。VSOP/Hv1は細胞外に存在する金属イオン、特に、亜鉛イオンに感受性があり、その結合部位がアミノ酸レベルで同定されています。亜鉛イオンはVSOP/Hv1が発現しているヒトの精子の受精機能に重要な役割を担っています。ヒトの精子は亜鉛イオン濃度が高い精液中に存在するためVSOP/Hv1の機能は抑制されています。しかし、射精後、女性の生殖器官内において亜鉛イオンは希釈されるためVSOP/Hv1の機能は脱抑制状態となります。そして、精子細胞内をアルカリ化することで精子の活性化を誘導します。
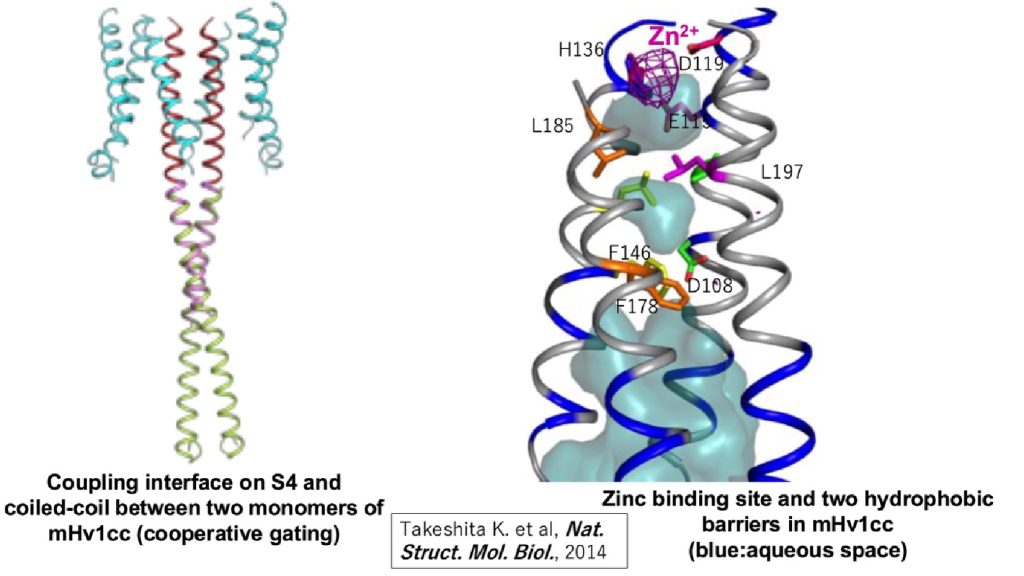
図3. 電位依存性プロトンチャネルの構造(mHv1cc: mouse VSOP/Hv1-CiVSP chimera) - 1. VSPにおいては、どのように電位センサーの働きにより酵素が活性化するのか?
- 2. VSOP/Hv1においては、どのように膜電位に応じてプロトン透過が起こるのか?
-
2. VSPの分子メカニズムと構造生物学
これまでに多くの研究から、チロシンホスファターゼやPTENががんや代謝障害の発症に重要な役割を果たすことが示されています。興味深いことに、VSPの細胞質酵素領域はこれらの酵素と構造的な類似点を持ちながら、膜電位の変化によってその酵素活性が制御されるという独自の特徴を有しています。この調節メカニズムを理解することは、チロシンホスファターゼやPTENががんやその関連疾患の発症にどのように関与しているかについて、貴重な洞察を提供する可能性があります。
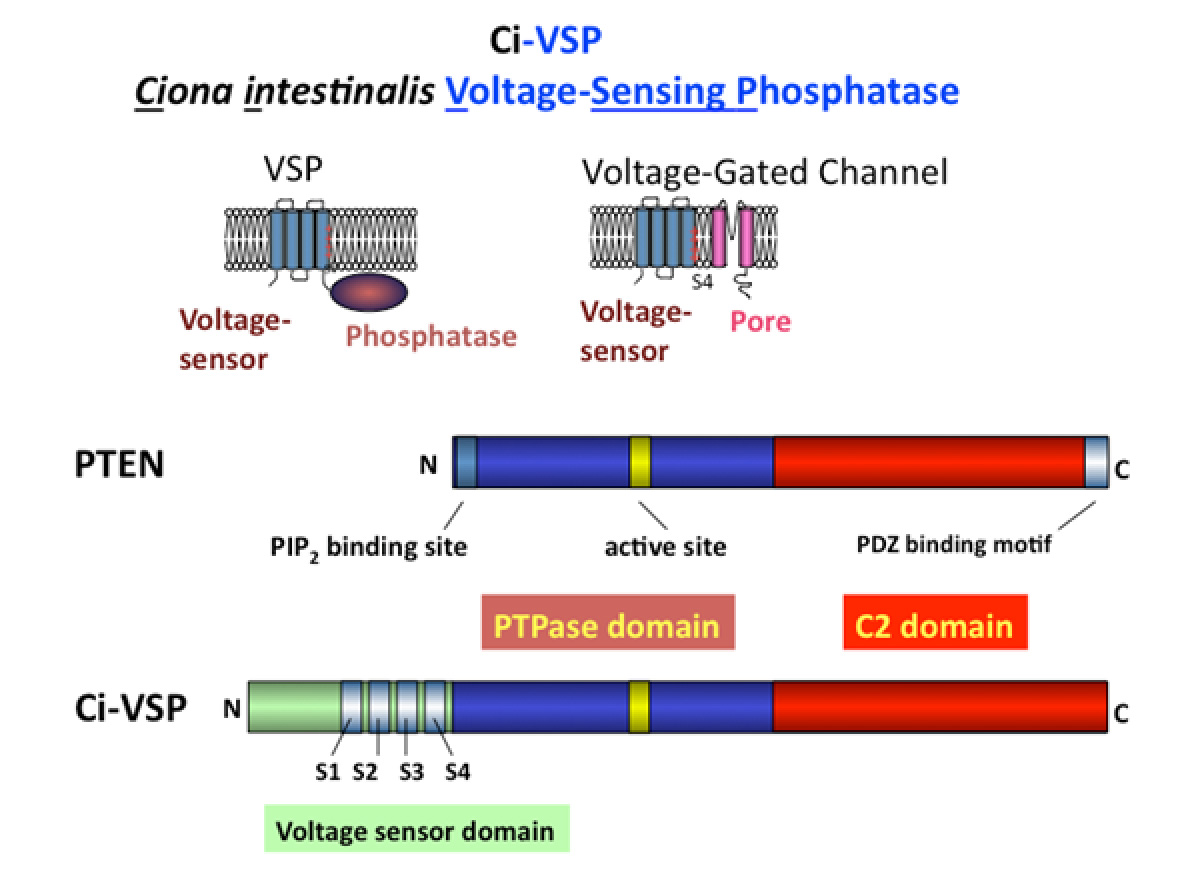
図4. VSPとPTEN VSPの特有の特徴は、膜電位の変化に応じて基質特異性を変化させる能力を持つことであり、この特徴は多くの酵素には見られません(Kurokawa Tら, PNAS, 2012)。電位センサーと酵素に関する私たちの研究により、この相互作用に関与する重要な領域が特定されました。ひとつは疎水性スパインであり、これは酵素が膜と結合する部位で、PTENやMTMR2を含む多くのホスホイノシタイドホスファターゼに保存されていることが判明しました(Kawanabe A.ら, eLife, 2018)。もうひとつの部位はS4セグメントの下部に位置し、膜電位を感知するために重要な正電荷を含んでいます。私たちは、これら二つの部位が膜の脱分極時に相互作用することを発見しました(Mizutani N.ら, PNAS, 2022)。特に、S4セグメントの下部は、PTENの酵素活性の調節に重要であることが知られているN末端と構造的に類似しています。これにより、VSPの電位センサーが、電位センサードメインを持たないPTENの酵素活性の固有の調節メカニズムを「乗っ取っている」ことが示唆されます。
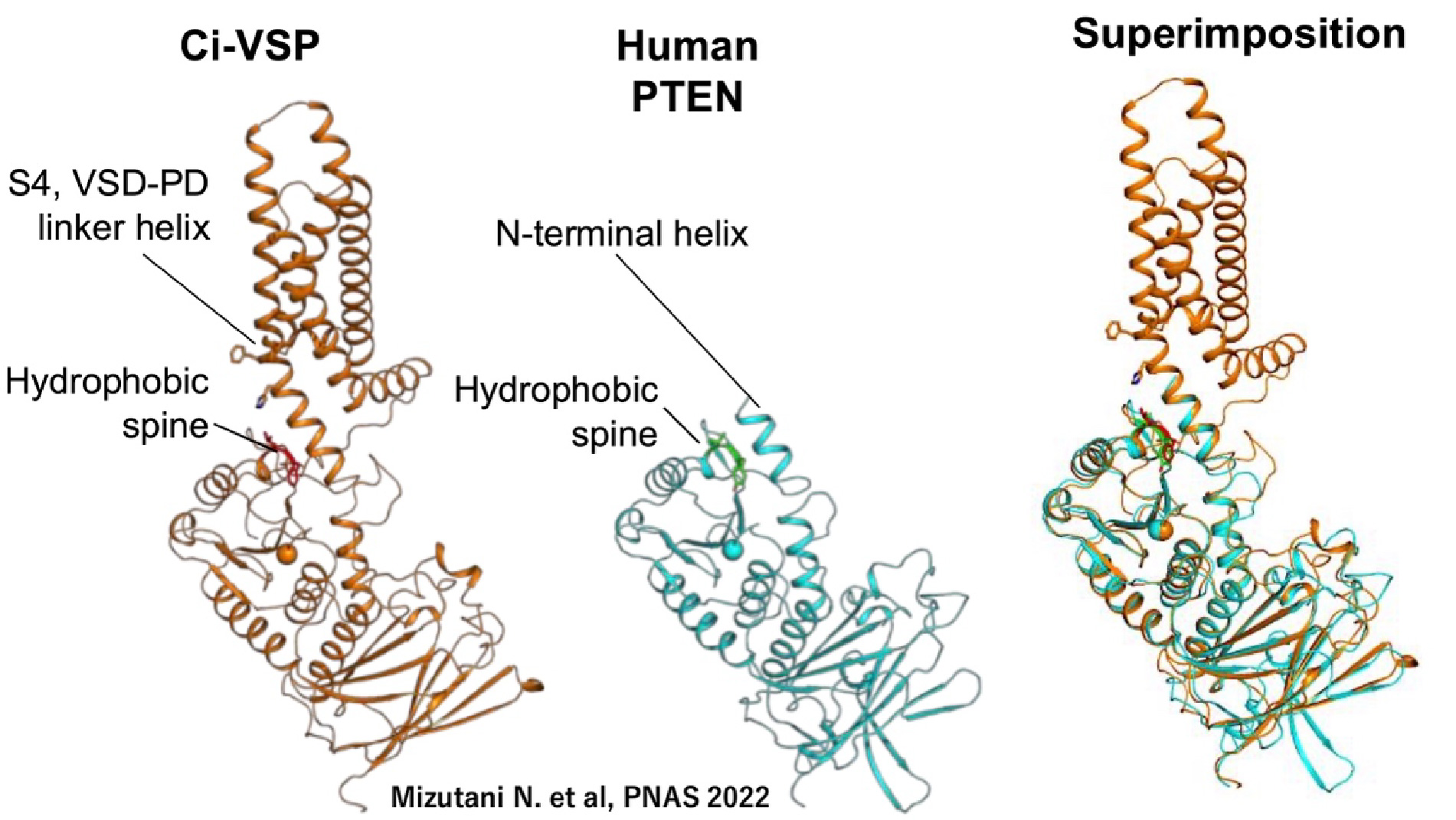
図5. VSPの機能に重要なドメイン間相互作用(Mizutani N.ら, PNAS, 2022) -
3. 血液細胞における電位依存性プロトンチャネルの役割
VSOP/Hv1(電位依存性プロトンチャネル=Hvチャネル)は海産無脊椎動物からヒトに到る様々な生物種に保存されており、哺乳類では、マクロファージや好中球といった貪食機能をもつ血球細胞に発現しています。プロトンを輸送する蛋白には、ATPを加水分解して細胞内からプロトンを運び出すプロトンポンプ(V-ATPase)がよく知られていますが、Hvチャネルは、プロトンポンプよりも短時間で大量のプロトンを排出することが可能です。膜電位の脱分極と細胞内外のpH差が引き金になってプロトン透過経路が開き、これによりプロトンイオン濃度勾配に従ってプロトンが通過するので、直接ATPを必要としません。
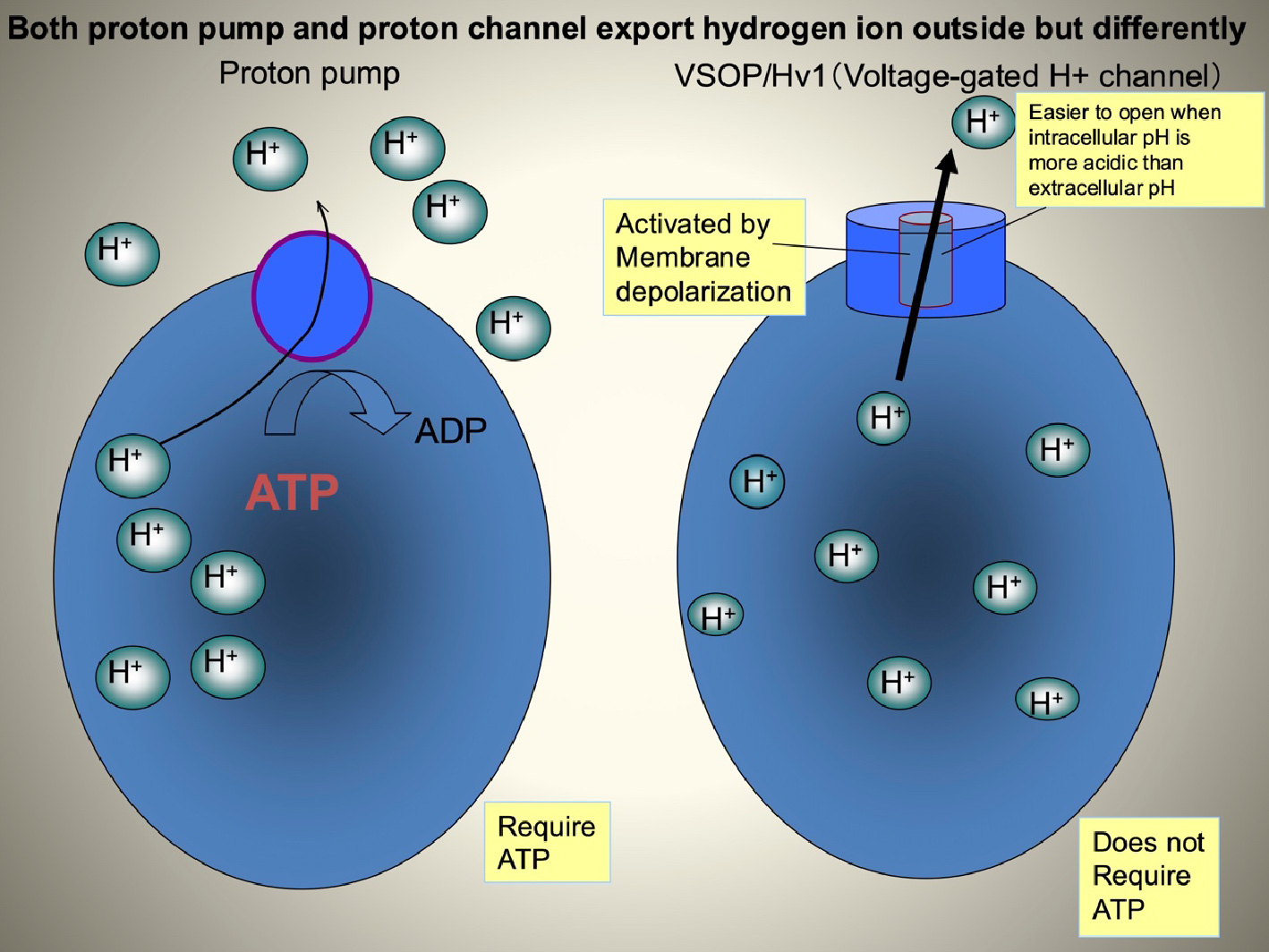
図6. 電位依存性プロトンチャネル(VSOP/Hv1)とプロトンポンプの違い これまでの電気生理学的な研究から、Hvチャネルは、貪食細胞において活性酸素の産生を促進することが示唆されてきました。最初に貪食細胞で活性酸素が作られる際には、酸素がO2-というマイナスの電荷をもつ物質として細胞の外へ放出されます。この状態が続くと電荷のアンバランスのため、細胞膜が大きく脱分極してしまい、オキシダーゼの活性が減弱してしまいO2-の産生が止まってしまいます。この電荷のアンバランスをキャンセルする役者としてプロトンを輸送する経路が想定され、Hvチャネルはまさにこの経路を担う実体として研究されてきました。私たちはVSOP/Hv1のノックアウトマウスを用いた解析を行い、プロトンの輸送により細胞質のpHと膜電位を制御することで、活性酸素の産生を助ける働きがあることを明らかにしました(Okochi et al, BBRC, 2009, El Chemaly et al, JEM, 2010、スイスのジュネーブ大学との共同研究)。VSOP/Hv1による活性酸素の制御は、その後、好中球の走化性、グルコース代謝、行動など予期しない表現型の発見につながりました (Okochi Y, et al, J. Leuk. Biol. 2020;Kawai T., et al, FASEB J. 2020、Dr. S. Kihara’s labとの共同研究;Kawai T., et al J. Neurochem, 2021、Dr. K. Sakimura, Dr. M. Abe, Dr. T. Miyakawaとの共同研究) 。注目すべきは、VSOP/Hv1ノックアウトマウスで見られる表現型が加齢と密接に関連していることです。現在、理化学研究所の荻沼博士との共同研究により、VSOP/Hv1と寿命の関連性を調べ始めています。私たちはまた地球科学、海洋科学の研究者と共同研究を行い、単細胞生物におけるVSOP/Hv1の役割を調べています(科研費:学術変革B「pH応答生物学」)。
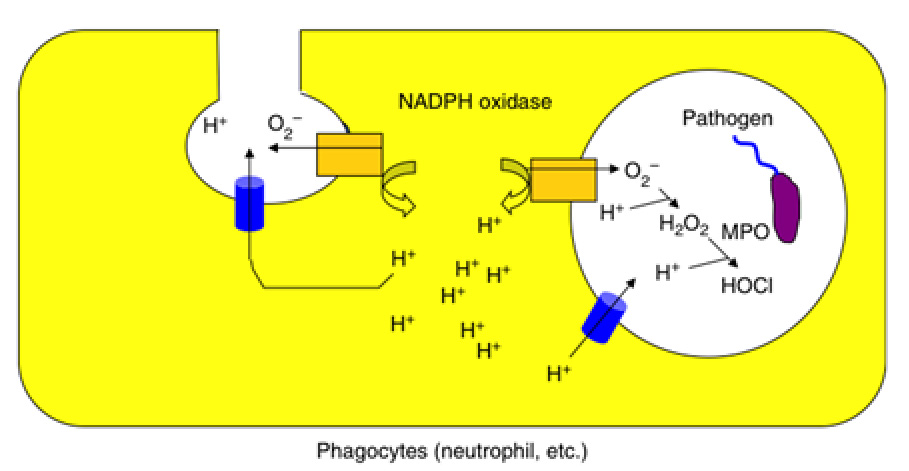
図7. 好中球およびマクロファージを含めた貪食細胞における電位依存性プロトンチャネル(VSOP/Hv1)の生理的役割 -
4. 膜電位の可視化にむけた分子ツール開発
神経回路の機能を理解する上で、個々のニューロンの活動を同時に計測することは極めて重要であり、また、電気生理学は、ガラス電極などが物理的にアプローチできない細胞や組織には適用することは困難で、膜電位の可視化は、こうした細胞での電気的現象の解明に有用であると考えられます。これまで有機化合物を用いた電位感受性色素やCa感受性色素がこれらの解析に用いられてきています。しかし、特定の細胞のみにプローブをターゲットさせることが困難であったり、シグナルの強度が強くないなどの問題があり、普遍的に用いられる手法はいままで確立されていませんでした。最近、複数のグループによりVSPの電位センサーに蛍光蛋白を結合させた分子が作成されました。タンパク性の膜電位プローブはGenetically encoded voltage indicators (GEVI)と呼ばれています。私たちの研究室のメンバーだった筒井助教は、理化学研究所の宮脇博士のグループに在籍時に、生理学研究所と協力して、蛍光強度変化の大きいプローブ(海産動物由来のタンパクを組み合わせたことから、Mermaidと命名)を作り出すことに成功し、培養細胞レベルで、ニューロンや心筋などの電気活動をイメージング化することに成功しました(Tsutsui et al., Nature Methods, 2008)。その後、マウスの大脳皮質の神経活動をモニターすることにも成功しました(Tsutsui T. et al., J. Physiol. 2013 PMID: 23836686)。
これまでに作成されたGEVIsのうち、約半数がVSPをベースにして作成されています。残りの半分は主にロドプシンをベースにしたGEVIsになります。VSPベースのGEVIsは特に多光子顕微鏡を用いたin vivo計測に向いているため、複雑な生物学的プロセスを理解するために行われる非侵襲的な神経活動の計測や他の細胞活動の計測において使用されています。
VSPはまた細胞膜に存在するPI(4,5)P2の量を操作する優れたツールとして使用されています。ツールとしてのVSPについて書かれた総説がいくつかありますので、ぜひご覧ください(Tsutsui H, Mizutani N, Okamura Y (2021) “Engineering voltage sensing phosphatase (VSP)”. Methods in Enzymology ISSN:0076-6879, 654:85-114; Okamura Y, Yoshioka D. “What voltage-sensing phosphatases can reveal about the mechanisms of ion channel regulation by phosphoinositides”. Biochem Soc Trans. 2023 Apr 26;51(2):827-839. PMID: 37052219.)。私たちは最近、VSPをツールとして使用することで、これまで見過ごされてきたトランスポーター(Mizutani N., Physiol. Rep. 2019)、リガンド依存性イオンチャネル(Okamura Y., 2024 Gordon Research Conference, keynote lecture)に対するPI(4,5)P2の役割を発見してきました。
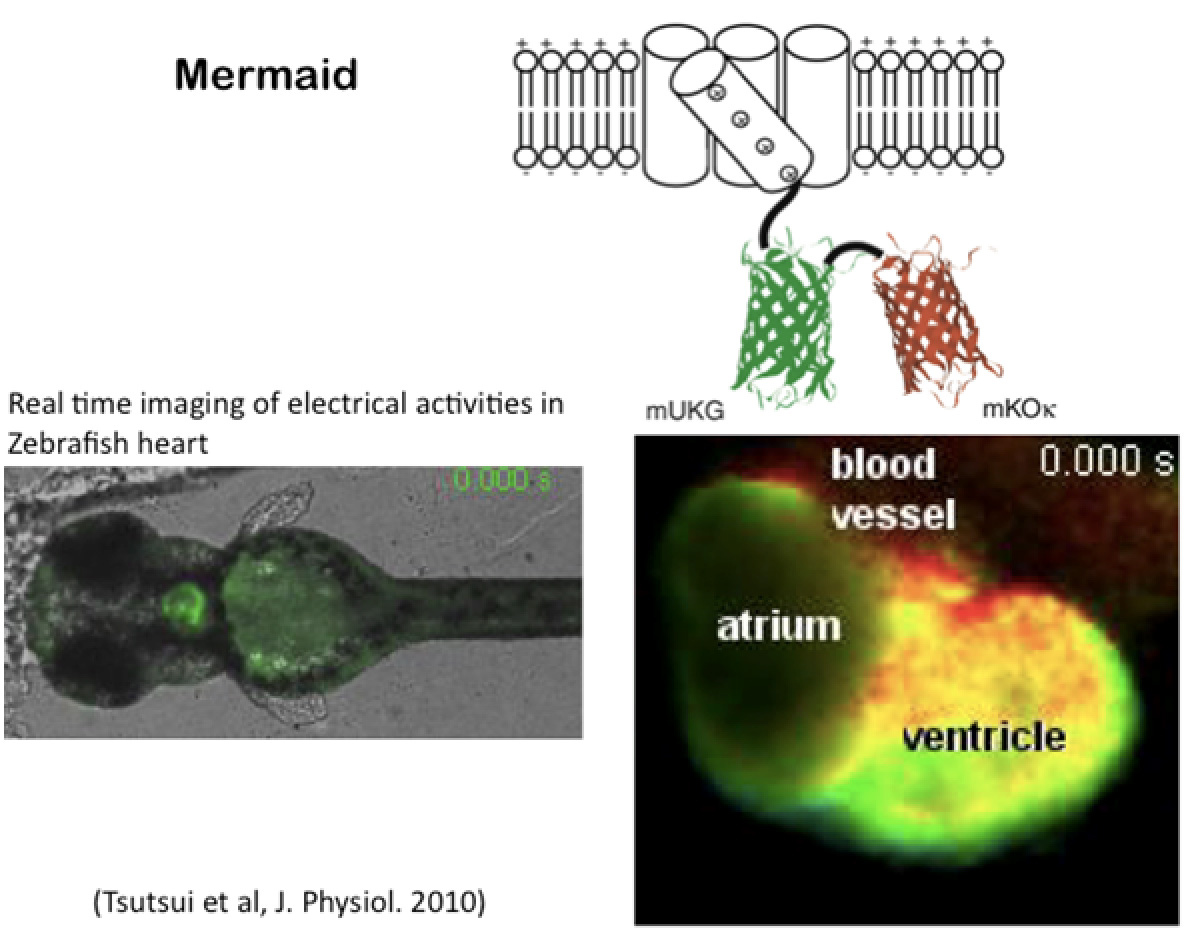
図8. VSPをベースに作製された膜電位を可視化するプローブの例。右下の画像は生きたままのゼブラフィッシュの心臓の膜電位変化をモニターした動画のスナップショット。 -
5. 生物現象における膜電位シグナルの新たな役割の解明
電位センサー分子に関する研究は、膜電位変化に関連した新しい分子経路の発見につながり、膜電位の生理学的役割を再評価する契機となりました。顕著な例として、受精時の多精子受精防止機構における膜電位の役割、左右軸決定への関与、非哺乳類脊椎動物における再生過程での機能が挙げられます。これらの過程の正確な分子機構は未だ不明ですが、膜電位変化の広範な生理学的影響を示しています。私たちは共同研究者と共に、VSP(Voltage-Sensing Phosphatase)や他の電位感受性タンパク質の生理学的役割を調査し、これらの複雑な過程をより深く理解するための研究を進めています。

図9. マウス精子の成熟過程におけるVSPの機能
電位センサードメインに単一アミノ酸変異を持つノックインマウスの解析により、VSPが未成熟な精子において膜電位を感知し、精子の機能に必要なイノシトールリン脂質環境を確立することが示されました(Kawai et al., Nature Commun, 2024)。VSPでは、膜電位の変化に応じて酵素活性が変化します。VSPは精巣で発現しており、精子の成熟に重要な役割を果たしています。図9は、分子特性に基づくVSPの生理学的役割を示しています。精子の成熟過程において、VSPは静止膜電位を感知し、酵素活性に依存してPI(4,5)P2を精子鞭毛全体に段階的に分布させます。このPI(4,5)P2の空間的分布は、PI(4,5)P2感受性の精子特異的カリウムチャネルであるSlo3の調節に不可欠であり、これがカルシウム流入の主要経路であるCatSperチャネルを介したカルシウム流入を調節します(Kawai et al., PNAS, 2019)。VSPの電位センサー変異体マウスを用いた私たちの最近の研究では、VSPの電位感知機能が、イノシトールリン脂質のプロファイルを調節することを通じて、精子機能成熟に重要であることが示されました。またこれにより、VSPが精子の成熟過程において膜電位を感知していることが実証さました。この研究は、哺乳類VSPオルソログが生理学的環境で電位感受性のPI(4,5)P2ホスファターゼとして機能することを初めて示しています(Kawai et al., Nature Commun, 2024)
-
6. 電位依存性イオンチャネルのターンオーバーと病態・老化との関連
神経細胞は、他の細胞と異なり、極性を持つ形態的特徴と生涯に渡って生き続けるという2つ特徴を持っています。神経細胞の機能が生涯を通してどのように維持されているか?という問いは重要な生物学的問題であり、この問題は脱髄や精神疾患、老化などの病態生理とも密接に関連しています。私たちは、神経細胞の機能に重要な役割を果たす軸索起始部(axon initial segment; AIS)とランビエ絞輪の2つの領域に局在するイオンチャネルのターンオーバーに着目して、研究を進めています。
AISは、哺乳類神経細胞の3つのコンパートメントから活動電位を計測した独創的な研究(J. S. Coombs, D. R. Curtis and j. C. Eccles (1957) The generation of impulses in motoneurones. J. Physiol. I39:232-249)によって初めて提唱された概念で、活動電位の開始点となる場所です。ランビエ絞輪は、有髄神経に特徴的な構造体で、ミエリン鞘の間に存在するミエリンのない1μm程度の間隙を指します。活動電位は、ランビエ絞輪において再発生または強化され、ミエリン間を“ジャンプ”することによって軸索を伝わっていきます(このメカニズムは、田崎一二先生が発見し、後に R. StampfliとA. Huxleyが確認した跳躍伝導(saltatory conduction)として知られています)。
AISとランビエ絞輪は構造と機能が似ており、電位依存性ナトリウムチャネル(Nav)・電位依存性カリウムチャネル(Kv)が局在し、共通した細胞骨格を有しています。これまでにこれら2つの構造体を形成する重要なタンパク質が同定されており、中でも特に重要なタンパク質として、NavやKvなどのタンパク質を係留する足場タンパク質アンキリンG (AnkG、遺伝子名Ank3)が知られています。AnkGは哺乳類に存在する3つのアンキリン(AnkG, R, B)のうちの1つで、Ank3遺伝子の変異は、双極性障害、統合失調症および自閉症と関連していることが知られています。
近年、AISの長さや位置がシナプス入力に応じて可塑的に変化することが明らかになりました(Kuba H., et al, Nature PMID;20543825 2010, etc)。この構造変化は、神経細胞自体の活動を恒常的に調節するためと考えられています。一方で、ランビエ絞輪はより静的な構造体だと考えられてきましたが、近年の研究から、ランビエ絞輪の長さも神経活動依存的に可塑的に変化する可能性が示唆されています。
私たちは、神経細胞のAIS・ランビエ絞輪に局在する電位依存性イオンチャネルが、生涯を通してどのように調節されているかを、空間解析と時間解析の2つのアプローチを用いて解析しています。一つ目のアプローチは、単一分子イメージングの手法を用いた電位依存性カリウムチャネルKCNQ2/3の三次元ナノスケールダイナミクスです(KCNQ2/3 はMチャネルとしても知られており、カエルの交感神経機能調節の研究において見出された)。二つ目のアプローチは、遺伝子組換えマウスを用いたアプローチで、蛍光タンパク質でイオンチャネルをラベルすることによって、古いタンパク質と新たに合成されたタンパク質をin vivoで可視化・区別する方法です。後者の遺伝子組み換えマウスを用いた研究は、新潟大学﨑村博士、阿部博士および福井大学深澤博士の協力の元、進めています。これら2つのアプローチを組み合わせて、私たちは、神経細胞の興奮性が生涯に渡ってどのように維持されるのか、そして、恒常性維持機構の破綻がどのようにして精神疾患を含む疾患を引き起こすのかを理解することを目指しています。
このプロジェクトは、「脊索動物の進化の過程で、どのようにして脊椎動物型の神経細胞に進化したのか?」という生物学と進化の根源的な疑問にも関連しています。私たちは、ホヤなどの尾索動物の神経生物学とイオンチャネルの研究を組み合わせて、この重要な疑問に光を当てたいと考えています(Okamura Y., et al, Neuron, 1994)。
-
7. 人工網膜による視覚機能回復の基礎的研究
網膜色素変性に代表される視細胞が変性する疾患では、視覚は永続的に失われ、その治療法はまだありません。このような患者の視機能を回復させる手段の1つとして、残存している網膜の神経細胞を電気刺激して視覚情報を入力する人工網膜があります。私たちは既存の方式よりも安全性に優れた新しい網膜刺激法である、脈絡膜上-経網膜刺激 suprachoroidal-transretinal stimulation (STS)方式の人工網膜を考案しました。そして電気生理学的方法や光学計測法を用いてSTS方式の人工網膜の機能を評価してきました。また、新しいデザインや刺激法を開発する基礎研究を行っています。人工網膜の研究開発には、医学、工学など複数の分野間の緊密な連携が重要であり、眼科学、生理学、工学、眼科医療機器会社の皆さんと共に研究を進めています。
![]()