私の研究履歴書
インターロイキン6発見物語:そして亜鉛シグナルへ
大阪大学生命機能研究科・医学系研究科、免疫発生学教室
理化学研究所免疫アレルギー科学総合研究センターサイトカイン制御グループ
平野俊夫
それは、今から20年以上前、1986年5月25日の日曜日、だれもいない研究室でオートラジオグラフィーのフィルムを現像すると、紛れもない、確かなスポットが目の前に忽然とあらわれた。インターロイキン6の精製を開始して、実に苦節8年、努力が報われた瞬間であった。
基礎医学への道:序曲はかくはじまりき(1972-1978年)
私が基礎医学の道を歩むことになったきっかけは、医学部一年生(大学3年)に近藤宗平教授の講義を聞いたことである。もともと、基礎医学者志望だったわけではない。しかし、暗記物の勉強に辟易していた学生時代、近藤先生の放射線基礎医学の授業を初めて聞いた瞬間に私の目の前の世界が急に開けた。近藤先生は京都大学で物理学を修めた後に医学生物学に転じた方で、物理学者らしい理路整然とした授業は、暗記に明け暮れた私にとって新鮮であった。早速その日から近藤先生の研究室に出入りして医学研究のイロハを教わった。1971年、卒業を間近に控えた私は将来のことを考え始め、免疫学の研究に進もうと決めた。その話を近藤先生に伝えると「平野君、それならすぐに米国に行きなさい」と言う。さすがに医師国家試験も受けずに「すぐに」行くわけにもいかないので、1972年3月に大阪大学医学部を卒業後、第三内科の山村雄一教授(元大阪大学総長、故人)の門をたたき、1年間臨床研修を受けてから米国に行くことにした。第3内科での最初の患者さんは、肺ガンの患者さんとSLEの患者さんで、文字どうり患者さんと一体となり戦ったが、刀折れ、矢尽き、臨床医学の限界を体のそこから実感した。この思いはその後羽曳野病院での勤務を通じてもますます大きくなった。また当時アメリカから帰国された渡邊 武先生に1年間免疫学のイロハを教えていただき、ますます免疫学への思いが強くなった。
1973年6月末に近藤教授が紹介して下さったボルチモアにあるNIHのDr. Makinodan(直接のボスはDr. Nordin)の研究室があるGerontology Research Center (現NIA)に留学した。現地では、渡邊先生に紹介していただいた第3内科の大先輩の岸本忠三先生(前大阪大学総長)が、自ら車を運転されてボルチモア空港に迎えに来て下さった。このようにして岸本先生とはボルチモアで初めてお会いすることになった(写真1)。さらに、高津聖志先生(現東大教授)、小里啓子先生(現NIH研究室長)ら年上の先輩方との出会いを経験した。そのご、研究分野も一緒だったこともあり、高津先生には公私ともに大変お世話になった。またジョンホプキンス大学の石坂公成先生と石坂照子先生にもお会いするという幸運に恵まれた。25歳、医学部を出たばかりでは、当然、研究成果は得られなかったが、良き先輩に囲まれて、研究の進め方と独立心だけはしっかりと心に刻んで3年後の1976年に第3内科に戻った。
当時の免疫学はといえば、1960年後半にT細胞、B細胞が発見され、免疫イムノグロブリンを中心とする免疫化学の時代から細胞免疫学へと流れは大きく歩み出した時だった。1971年にワシントンで第1回国際免疫学会が開催され、DuttonがT細胞のかわりをする液性因子が存在することを発表、引き続き1972年にSchimpl と Weckerが T cell replacing factor (TRF)の存在をNatureに発表して注目を集めていた。ちなみに1971年に山村先生を中心として日本免疫学会が結成された。古くは1944年のMenkinによる内因性発熱因子、1953年のLevi-MontalciniらによるNerve Growth Factor、1965年のCohenらによるEpidermal Growth Facotr の存在の発見(Cohen and Levi-Montalcini,1986年ノーベル医学生理学賞)、1954年の長野、小島によるウイルス干渉現象の発見(インターフェロン存在の発見)、1965年のKasakura and LowensteinによるLymphocyte blastogenic factor、1966年のMigration inhibitory factor, 1967年のLymphotoxin、1976年のT cell Growth Factor (TCGF) の存在の発見など、各種活性を有する液性因子が存在することの発見が相次ぎ、1978年当時には、200種類以上の液性因子の存在が報告されていた。もちろんこれらの液性因子はすべて微量分子であり、その実体は全く不明だった。T細胞の抗原受容体も不明であり、その実体解明に向けて華々しい競争が繰り広げられていた。1975年にKohler and Milstein(1984年ノーベル医学生理学賞)によりモノクロナル抗体作成の報告がNatureに掲載されるとともに、1976年にはTonegawaらにより免疫グロブリン遺伝子再構成が発見された(1987年ノーベル医学生理学賞)。大地震のまえの地底でのエネルギーの蓄積のごとく、エネルギーの蓄積が爆発、あるいは爆発寸前、ダイナミックな現代免疫学がまさにスタートしようとしている時だった。
私の歩んできた研究者人生を、愛好しているブルックナーの第7交響曲になぞらえるとこの時期は、私自身の交響曲はまだ始まらない、あえて言うなら序曲と言うことになる。かくして序曲は、アダージヨ、非常にゆっくりとしたテンポではじまった。多くの若者がそうであるように、少し不安はあっても、夢と希望にあふれる若者が一人、ゆっくりと世界に歩み出した。
第一楽章(アレグロ・モデラート)1978-1989、インターロイキン6の発見
第一楽章は運命的に幕を開いた。羽曳野病院での結核性胸膜炎患者との運命的な出会いが、インターロイキン6の発見へと私を導いた。
帰国後2年間大阪大学第三内科で働いた後、1978年に大阪府立羽曳野病院に出ることになった。現在は一般病床が1000床もある総合病院だが、当時の羽曳野病院はまだ結核療養所の名残りがあり、患者さんも、結核や呼吸器疾患、さらにアレルギー疾患の方がほとんどだった。羽曳野病院での私の上司は露口泉夫先生(元・羽曳野病院院長)だった。露口先生からは患者管理の手法をいろいろ教わったが、「胸膜炎の患者の胸水にはTリンパ球が非常に多い」という話は私の好奇心をいたく刺激した。胸膜炎の患者さんからは治療のため、1L近い胸水を抜くことも多々あった。胸水1Lの中に10億このリンパ球が存在している。そんな患者さんが病院には何十人といた。しかも胸水リンパ球を結核菌体成分で刺激すると、その培養上清中には非常に強い抗体産生誘導活性があった。当時はサイトカインの存在は知られていたが、その本体は全く不明だった。サイトカインを精製していた研究者は世界でも数人だったが、なんとか精製ができないかと考えた。同僚の寺西強医博と日中は受け持ちの患者さんの治療、夜はIL-6の精製をしていた頃、当時大阪大学の総長の山村先生から「明日、阪大に来るように」と電話があった。総長室に行くと「平野君、学校の先生にならんか?」と言われた。九大から熊本大学に行かれた尾上薫教授が、助教授を探しているとのことで私に白羽の矢が立ったようだ。熊本大学では引き続きIL-6の精製とその解析を4年間かけて行い、等電点5.1で分子量2.2万のB細胞分化因子(TRF様因子/BCDF)、すなわちIL-6の存在を発見し、その精製を試みるとともに、作用機作の研究を続けた(1)。
1979年から80年にかけて、谷口維紹氏によるインターフェロンベーター、Shigekazu Nagata,Charles Weissmannらによる インターフェロンアルファーの遺伝子クローニングの成功というサイトカイン研究史におけるマイルストーンといえる事件があった。当時の私にはインターフェロンは免疫とは関係ない遠い世界の出来事に思えたが、1983年の谷口維紹氏によるIL-2遺伝子のクローニング成功の大ニュースを新聞で知り、少なからず衝撃を覚えるとともに、私と同世代の若い研究者の存在に深い敬意を評するとともに、いつかは追いつきたいという思いが沸いてきた。そして、遺伝子工学の手法も取り入れるべきだろうかと考えていたころ、岸本忠三先生が阪大に新設された細胞工学センターの教授になり、私に助教授として一緒に研究をやらないかと誘っていただいた。細胞工学センターに移ったのは1984年1月、そのころには私の研究も胸水細胞や扁桃腺細胞から精製する手法からウイルスでトランスフォームしたT細胞からサイトカインの同定をする手法に移っていた。1984年12月、蛋白研の綱澤先生らの助けを借りてN端の部分的アミノ酸配列の決定に成功した。「これで早晩、遺伝子の単離も成功する」と思ったが、1年経っても期待した成果は出ず、1985年の年末には、いら立ちとストレスから不整脈が頻発し夜も眠ることができず、研究者の道をあきらめようかと思った。正月明けに、友達の循環器専門医に診察を受け、心因性の不整脈であることがわかった。こんなことで、人生を棒に振るのはばかばかしいと思い直し、もう一度白紙の心でIL-6の精製を一からやりなおした。今から思えば、このときは山の頂上の直下、息を切らしながら頂上を目差す登山者のそれであった。頂上が目の前にあることはまだわからない。息絶え絶えの状態であった。その時頂上は突如目の前に出現した。1986年5月25日、日曜日の午前11時に研究室に来てみると、三つの異なるプローブと結合している遺伝子が確認できた(写真2)。羽曳野病院で精製を開始してからじつに苦節8年、ついにIL-6遺伝子の単離に成功した瞬間だった。その日の午後、一緒に苦楽を共にしていた保川清君(現京都大学助教授)、渡部保夫君(現愛媛大学助教授)、松田正君(現北海道大学薬学部教授)らと大学の近くの喫茶店で興奮しながら今後の実験計画を立てたのがつい昨日の様に思える。幸いにも我々の 研究成果は1986年11月にNature誌に掲載された(2)。同じ年の9月に26kDa 蛋白の、10月にはインターフェロンベーター2のクローニングの報告が、Eur. J. Biochemistry とEMBO J にそれぞれ掲載された。驚いたことにこれらの分子はすべて同じ構造をしていることが判明した。あと1ヶ月論文投稿が遅れていれば、我々の論文はNature に掲載されなかっただろう。1986年の夏にカナダのトロントで開催された国際免疫学会に予定どうり参加した。学会にはほとんど出席せずにホテルに閉じこもり論文執筆をしなかったらどうなっていたことか!研究における競争という厳しい現実を心の底から実感した瞬間でもあった。 その後、ミエローマプラズマサイトーマ増殖因子や、肝細胞刺激因子など種々の分子は、すべて我々がクローニングした分子と同じものであることが明らかになった。各々のグループが異なる名称を使用していたので、1988年のニューヨーク・アカデミーの主催する国際会議においてインターロイキン6という名称に統一された(写真3、4)(1)。引き続き、大学院生の田賀哲也君(現熊本大学教授)、山崎勝彦君(現厚生労働省)、日比正彦君(現理化学研究所発生再生科学総合研究センターチームリーダー)らとレセプターの構造を決定、インターロイキン6の全貌を解明することができた。1992年にハンガリーのブタペストで開催された第8回国際免疫学会で岸本先生とサンド免疫学賞(現在のノバルティス免疫学賞)を共同受賞する感激を味わうことができた(写真5)。夜のドナウ川に映る街灯の美しさが星のきらめきのごとく脳裏に焼き付いている。1978年から1989年の約10年間はまさに分子生物学の開花とともに、第1楽章は将来への暗示をともない華々しく展開、1989年11月に大阪大学医学部教授に就任し、その幕を閉じた。この間、当時細胞工学センターの教授をされていた谷口維紹先生にはいろいろ教えていただき、公私ともに大変お世話になった。大学近くのマンションへの夜道を谷口先生と語りながら歩いたのが懐かしく思い出される。
第二楽章(アダージヨ、非常に荘厳に、かつゆっくりと)1990-2000、インターロイキン6シグナル伝達機構の研究
第二楽章は大阪大学医学部附属バイオメディカル教育研究センター腫瘍病理教室で始まった。
長い道のりの果て ついにいま、確かな一歩を
果てしのない道が再び目の前に
そのはるか彼方に 希望の星が静かに輝いている
踏み出そう、勇気をもって 新たな第一歩を
研究室は大阪市の中心部の中ノ島にある私が医学部教育を受けた懐かしい旧阪大医学部にある旧蛋白研の古い建物の中にあった。1年後には吹田の新しい医学部キャンパスに移転の予定のため、古い建物にある研究室は、雨漏りすれども、抜本的な修理は行われず、応急的に天井にブリキ缶を吊るし、ゴム管でたまった雨水を窓の外に誘導するというものだった。かくして我が研究室は雨ともなると優雅な雨垂れの音楽が響き渡るという状態だった。それでも窓の外に咲き誇る満開の桜の花は目に染みた。
遠心器音なり出せば心打ち眼にしむ桜あでやかに舞う
中嶋弘一君(現大阪市立大学医学部教授)、松田正君、改正恒康君(現理化学研究所免疫アレルギー科学総合研究センターチームリーダー)らと、インターロイキン6の作用機構の解明に向けての非常に地味な、しかし重要な研究を開始した。IL-6受容体を介して少なくとも2つのシグナル伝達、すなわち、STAT3を介するシグナル伝達経路と、SHP2/GAB/MAPKを介するシグナル伝達経路が活性化されることを証明するとともに、それぞれのシグナル伝達経路が細胞の増殖や、分化、生存、細胞運動にどのように関与しているかを明らかにした(3,4)。さらに、これらのシグナルが生体内でどのような役割を果たしているかを明らかにするために、gp130を介するそれぞれのシグナルのみを特異的に欠失した変異gp130を発現しているマウスの作成を試みた。そして2000年、大谷卓也君(アメリカ留学中)、日比正彦君らとレセプターの点変異マウスの作成に成功、試験官内のみならず、生体内でのインターロイキン6の作用機構の解明に成功した(5)。単なる遺伝子のノックアウトの手法では解明することが困難であるとされていた、特定のサイトカイン受容体を介する、特定のシグナルの生体内での役割を明らかにすることができた。この間、山中庸次郎君(カナダ留学中)、伊藤素行君(現名古屋大学特任助教授)、深田俊幸君(現理化学研究所免疫アレルギー科学総合研究センターサブチームリーダー)、西田圭吾君(現理化学研究所免疫アレルギー科学総合研究センターサブチームリーダー)、藤谷与士夫君(現順天堂大学医学部助教授)、大谷卓也君ら多くの優秀な大学院生が育っていくという、幸福感を味わうことができた。
第三楽章(スケルツォ、非常に速く)2000-現在、インターロイキン6が自己免疫疾患などの病気に関与していることの発見とその機構解明
第三楽章は石原克彦君(現川崎医科大学免疫学教授)、熱海徹君(アメリカNIH留学中)らによる、インターロイキン6のレセプターの点変異により関節リウマチのような自己免疫疾患が発症するという発見により幕がひらいた。
関節リウマチなどの自己免疫疾患は複数の遺伝要因と環境要因により発症する難病で、その原因はいまだ不明だ。1987-88年にかけて、心房内粘液腫の患者に観られる自己免疫様症状が、粘液腫を摘除することによって消失することに注目し、粘液腫が産生する何らかの因子が関与している可能性を考え、その因子の本態がIL-6であることを明らかにした。また、関節リウマチ患者関節液中には、IL-6が著増していることを見つけ、IL-6が自己免疫疾患に関与しているのではないかと考えていたが、決定打はなかなか得ることはできなかった(1)。いろいろモデルを考え、IL-6と関節リウマチのような自己免疫疾患の関係を明らかにする努力を模索しつつも失敗の連続で、このことは一日とも私の脳裏を離れることはなかった。そんなとき、2000年に石原君、熱海君らが、上述したSHP2シグナルを欠損している変異gp130を発現しているノックインマウス(gp130F759マウス)は、gp130を介するSTAT3シグナルが亢進しているとともに、加齢により自己抗体を産生し関節リウマチ類似の関節炎を発症することを見つけた。最初に報告を受けたときには、にわかには信じられなかった。確かにIL-6シグナルが異常になっているgp130F759マウスは、老化とともに、例えばプラズマ細胞腫のような何らかのがん性病変をきたすことは期待してはいたが、関節リウマチのような複数の遺伝子や環境要因がからんでいる自己免疫疾患がIL-6 の単独の異常だけで自然発症することは、願望であっても、考えがたかった。しかし、まさに“事実は小説より奇なり“、まぎれもなくIL-6シグナルルの異常で関節リウマチ様の関節炎が自然発症した。興奮して眠れない日々が続いた。ついに、IL-6などのサイトカイン受容体シグナル異常により自己免疫疾患を発症することを証明した(6)。サイトカインと自己免疫疾患発症の機構を考える上で、非常に重要な知見がもたらされた。IL-6受容体に対する抗体療法が関節リウマチに効果があることが明らかにされ、臨床応用がまじかになりつつある今、我々の発見により、なぜ抗IL-6受容体抗体が関節リウマチに効果があるかを科学的に示したのみならず、なぜIL-6の異常で関節リウマチなどの自己免疫疾患が発症するかの機序を解明する道が開けた。第2楽章の静けさとは一転、スケルツォへ、研究は興奮の渦の中に急展開を見せ、現在、村上正晃君(現大阪大学生命機能研究科助教授)という力強い共同研究者を迎えて、第3楽章はインターロイキン6の異常でなぜ自己免疫疾患が発症するかの仕組みの解明に向かって展開中だ。そして、2006年には澤新一郎君(現在フランス留学中)と村上君により、免疫システムと非免疫システムの相互作用により関節リウマチのような自己免疫疾患が発症するのではないかというモデルを証明することができた(7)。
第四楽章(フィナーレ、活発に、しかし速すぎず)1998-現在、亜鉛は細胞内シグナル伝達分子としての作用を有するか?
さらに第3楽章の進行と平行して第四楽章がスタート、亜鉛シグナルの世界への新たな道が忽然と目の前に現れた。
吹田の地我がふみ込めし足跡に新たなる一歩こころは踊る
まず1998年に山中君(カナダ留学中)、日比君らが、ゼブラフイシュのシュぺーマンオーガナイザー形成に必須のホメオボックス蛋白であるダルマを発見した。1924年にHans SpemannとHilde Mangoldにより発見されたオーガナイザーは、体が形成される形づくりの過程(ボディープラン)において、細胞分化と細胞運動を制御している。人間をはじめ、生き物の<かたちつくり>のために非常に重要な過程であり、オーガナイザーにより誘導される原腸陥入における細胞運動が、IL-6のシグナル伝達分子であるSTAT3により制御されていることを、山下晋君(現大阪大学生命機能研究科学内講師)、宮城智恵美さん(現大阪大学医学系研究科特任研究員)らが、2002年に発見した(8)。さらに、その仕組みに、私たちが生きていくためには欠かせない金属である亜鉛の細胞内含量を制御している膜蛋白、亜鉛トランスポーターLIV1/Zip6が関与していることを発見した(9)。すなわちSTAT3の標的遺伝子としてLIV1を同定し、亜鉛トランスポーターLIV1がZincフィンガー転写因子スネールの核への局在に必要であることを明らかにした。LIV1はこれまで臨床統計学的に乳癌の転移との関係が報告されていたが、その生体内における機能は未知だった。我々はサイトカイン研究から発生学の世界へと導かれ、思いもよらず亜鉛の世界に遭遇するに至った。亜鉛欠乏は免疫不全、成長障害、味覚障害、神経系の異常など、さまざまな障害を引き起こすことが知られている。事実、亜鉛要求性の蛋白は300種、亜鉛要求性の転写因子も多数存在する。しかしながら亜鉛はこれら酵素や転写因子の酵素活性や高次構造を保つために構成的に必要な必須得微量元素と考えられていた。亜鉛が神経末端から分泌され神経伝達分子(neurotransmitter)として作用する可能性は報告されているが、いわゆる細胞内セカンドメッセンジャーとしてシグナルル伝達に関与している報告はない。亜鉛トランスポータであるLiv1/Zip6が亜鉛結合サイトを有した転写因子Snail の細胞内局在を規定することに関与している事実は亜鉛が細胞内シグナルル伝達分子としての役割を担っている可能性を強く示唆している。さらに2006年に、免疫応答開始の重要な過程の一つである樹状細胞の成熟活性化の過程で樹状細胞内の遊離亜鉛濃度が減少すること、このことが亜鉛トランスポーターの発現制御により行われていること、さらに遊離亜鉛濃度の減少が樹状細胞の成熟活性化に必須の過程であることを北村秀光君(現北海道大学助教授)と村上君らが見いだした(10)。この事実は亜鉛が免疫応答における細胞内シグナル伝達分子として作用しうることを強く示唆している。亜鉛が細胞内シグナル分子として作用しることや、サイトカインシグナルと亜鉛シグナルがリンクすることの発見は、免疫、炎症、発生、癌など領域を超えて生体の恒常性維持機構の研究に大きなインパクトを与えることが予想される。第4楽章に至り、我々のインターロイキン6の研究は、免疫学を超えて、生命科学の基本原理の解明へと、無限の可能性を秘めて進行中である。
目の前の山を登りきることが重要
研究室の若い人たちに例え話として山登りの話しをする。登山家は「どうせ登るのなら高い山に登りたい」と考える。しかし、私たち研究者にとっては、山が高いか低いかは登ってみないことには分からない。斬新な研究だと思っていたものがつまらなかったり、途中で投げ出したくなったりするかもしれない。しかし、「目の前の山を登りきる」ことが重要だ。
山登りをした人ならお分かりでしょうが、頂上を目指して歩いているときは、頂上では、どのような景色が展望されるのは全くわからない。山の頂上に登り切って初めて目の前に新たな景色が広がる。頂上にたって初めて自分が登った山の高さがわかる。その山は予想に反して低いかもしれないし、高いかもしれない。たとえその山が低くても頂上に立てば、目の前に素晴らしい高山がそびえているかもしれない。新たな予期しなかった秀峰がそびえているかもしれない。次にめざす山が見えるはずだ。頂上近くなるとたいていはきつくなりあきらめようとする。頂上近くが、登山において最もしんどい時だ。脱落の危険性が最もあるときだ。中途半端でいくつもの研究を投げ出すと、いつまでも中途半端な研究者にしかなれない。たとえ100回山を登っても、一度も頂上に立ったことがない人は、一度だけ登山して頂上にたった人には決して及ばない。たとえ低い山でも頂上に立つことができた人のみが新しい景色を見ることが、更なる展開をつかむことができる。これは研究だけではなく、我々の人生すべてに共通する事だ。
IL-6がB細胞に作用して抗体産生を誘導する以外にも肝臓の細胞に作用して急性期蛋白の産生を誘導したり、多発性骨髄腫の増殖因子であるというのは、予想していなかった事実だ。1988年に関節リウマチの患者さんの関節液中にIL-6が多量存在していることを見いだし、IL-6が関節リウマチの病態に関係しているのではないかということが想像されたが、このようなことは、研究開始時点では想像すら難しく、研究の結果、頂上まで登って初めてわかったことだ。IL-6が関節リウマチ関節液中に多量存在しているからといってもIL-6がリウマチの原因なのか、それとも副次的なものかはわからない。地道な研究の結果、IL-6受容体のある部位にミューテーションを入れたマウスは加齢により自然に関節リウマチ様関節炎になることがわかった。IL-6のシグナル異常により関節リウマチが発症するということを証明できた。さらにこの機序を解明すれば関節リウマチなどの自己免疫疾患の発病メカニズムがわかるはずだ。今年で研究者生活34年目となったが、IL-6シグナル異常で関節リウマチのような自己免疫疾患が発症することを証明できたことは研究者冥利につきない。1972年医学部卒業当時の"夢"が“IL-6の発見とIL-6シグナル異常で自己免疫疾患が発症することを明らかにした”ことにより現実のものとなった。しかし科学に終わりはない。IL-6の研究が亜鉛シグナルという全く予想しなかった新しい世界へと私を導いてくれた。そこには自然の奥深さを感じる。
山村雄一先生が1990年6月に亡くなられる2ヶ月前の4月に先生からいただいた色紙がいつも私の目の前にある(写真6)。
樹はいくら伸びても 天まで届かない
それでも伸びよ 天を目指して
最後に多くの良き共同研究者と山村雄一先生や岸本忠三先生をはじめとする多くの指導者に恵まれたこと、家族の理解があったこと、多くの幸運に恵まれたこと、今日、この瞬間まで研究を続けることができたことを、ただただ感謝するのみ。“天の時、地の利、人の和”が私を今へ導いてくれたことを心の底から思うのみ。
文献
囲み記事
インターロイキンとは
インターロイキン6などのサイトカインは細胞の増殖、分化、生存、運動などを制御することにより、免疫応答、造血反応、炎症反応などを制御している生理活性分子である。例えば、抗原刺激を受けたT細胞やB細胞がキラーT細胞や抗体産生細胞などのエフェクター細胞へ増殖、分化するためには一連のサイトカインが重要な役割を果たしているし、ワクチン接種の結果免疫記憶が誘導され維持されるためにもサイトカインは重要な役割を演じている。赤血球や好中球が作られるためにはエリスロポエチンやG-CSFが必要である。現在では種々のサイトカインの存在が明らかになっているが、1970年代においては、インターロイキン6(IL-6)などのサイトカインの本態は全く不明であった。1979―80年のインターフェロンa/bのクローニングに始まり、1980年代には我々のIL-6を含めて代表的なサイトカインが次々とクローニングされその構造が明らかになった。引き続き1980年代後半には、サイトカイン受容体の構造明らかにされた。その結果、サイトカインの特徴として、1つのサイトカインが様々な機能を持つこと(多機能性)、ファミリーを形成する複数のサイトカインが共通の作用を示すこと(冗長性)が明らかとなり、広義には、増殖因子、造血因子、ケモカインなども含まれようになった。また、リガンド特異的に結合するサブユニットとファミリーサイトカインで共通に使われる信号伝達のサブユニットからなるサイトカイン受容体の基本構造が解明された。1990年に入り、信号伝達経路が明らかにされていった。このような機能をもつサイトカインの重要性は臨床医学、基礎医学、基礎生物学的研究による知見の蓄積により確固たるものとなった。さらにトランスジェニックマウスやノックアウトマウスなどの発生工学的な解析によりサイトカインの生体における役割が明らかになった。特に発生工学的に樹立された疾患モデルマウスの樹立は、様々な難病の病因・病態の理解に貢献し、今やサイトカインの作用を人為的に制御することにより難治疾患を根治する可能性が大きく展開している。さらに、エリスロポエチン、G-CSF、インターフェロンなどのサイトカインはすでに臨床医学において大きな成果を上げている。またTNFに対するモノクローナル抗体が関節リウマチなどに効果があることがわかり臨床の場で成果を挙げている。引き続きインターロイキン6の受容体に対するモノクローナル抗体も臨床応用されつつあり、今後の展開が大いに期待されている。
著者プロフィール
平野 俊夫(ひらのとしお)
昭和47年大阪大学医学部卒業、昭和47年大阪大学医学部第三内科入局(山村雄一教授)、昭和48~51年アメリカNIH留学、昭和53~55年大阪府立羽曳野病院内科医員、昭和55~59年熊本大学助教授(医学部附属免疫医学研究施設生化学部門)、昭和59~平成元年大阪大学助教授(細胞工学センター)、平成元年11月大阪大学教授(医学部附属バイオメディカル教育研究センター腫瘍病理)(平成11年3月まで)、平成9年10月大阪大学医学部附属バイオメディカル教育研究センター長(平成11年10月まで)、平成9年10月大阪大学評議員(平成11年10月まで)、平成11年4月大阪大学教授(大学院医学系研究科附属バイオメディカル教育研究センター)(平成13年3月まで)、平成13年4月大阪大学教授(大学院医学系研究科病理病態学講座)(平成14年4月より兼任、現在に至る)、平成13年9月理化学研究所 免疫・アレルギー科学総合研究センター グループディレクター兼任(現在に至る)、平成14年4月 大阪大学教授(大学院生命機能研究科免疫発生学)(現在に至る)、平成15年4月大阪大学評議員(平成18年3月31日まで)、平成16年4月大阪大学大学院生命機能研究科長(平成18年3月31日まで)、日本免疫学会会長(平成17年1月―平成18年9月)、平成18年3月 日本学術会議連携会員(現在に至る)、第36回日本免疫学会学術集会会長(平成18年)
所属学会:日本免疫学会、アメリカ免疫学会(昭和55年~)、日本癌学会、日本生化学会、日本分子生物学会
専門分野:免疫学
受賞:ベルツ賞(昭和61年)、日本チバガイギーリウマチ賞(平成2年)、The Sandoz Prize for Immunology(平成4年)、大阪科学賞(平成9年)、持田記念学術賞(平成10年)、ISI 引用最高栄誉賞(ISI Citation Laureate Award, 1981-1998)(平成12年)、藤原賞(平成16年)、日本医師会医学賞(平成17年)、紫綬褒章(平成18年4月) 、クラフォード賞 (平成21年1月)
写真1、岸本先生夫妻、高津先生夫妻らと、
岸本忠三先生夫妻(右側2人)、高津聖志先生夫妻(左端、左から三人目)、とともに(左から二人めが本人)。アメリカ ボルチモア、1974年
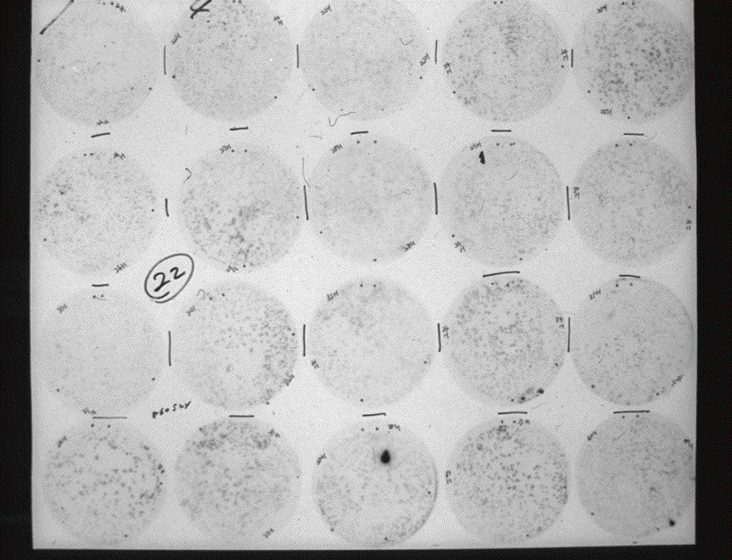
写真2 インターロイキン6遺伝子の発見
ヒトインターロイキン6をコードするcDNAクローンを初めて同定したオートラジオグラフイー

写真3 インターロイキン6の名前が世に出る。
1988年12月12日―14日にニューヨークにて、ニューヨークアカデミー主催の国際会議 ”Regulation of the Acute Phase and Immune Responses:New Cytokine” が開催された。この会議中に、interferon beta2, 26kDa protein, B cell stimulatory factor 2 など、種々の名前で呼称されていた分子をインターロイキン6と呼称することが決定された。

写真4 ニューヨークアカデミーサイエンス年報表紙
1988年12月12日―14日にニューヨークにて、ニューヨークアカデミー主催の国際会議 ”Regulation of the Acute Phase and Immune Responses:New Cytokine” が開催され、世界中からこの分子の研究に関与していた科学者が初めて一同に会した。この会議の内容が記載されたAnnals of the New York Academy of Sciences vol. 557, 1989, ”Regulation of the Acute Phase and Immune Responses:Interleukin-6”の表紙である。

写真5 インターロイキン6の研究で岸本忠三先生とサンド免疫学賞を共同受賞
岸本先生、ハーバード大学のストロミンジャー教授(MHCの研究でサンド免疫学賞を受賞)と記念写真
1992年8月、第8回国際免疫学会 ブタペスト、ハンガリーにて

写真6 樹はいくら伸びても 天まで届かない それでも伸びよ 天を目ざして 1990年 山村雄一
|
|
|