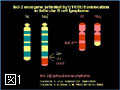
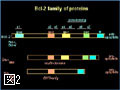
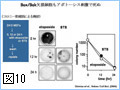
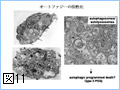
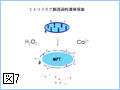
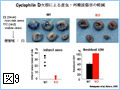
Bcl-2ファミリーのアポトーシスメンバー促進メンバーの多くは、生細胞中ではミトコンドリア外に存在し、アポトーシス刺激により活性化されミトコンドリアに移行する(図4)。また、Bcl-2ファミリーに属さず、アポトーシス刺激に応じてミトコンドリアに移行し、ミトコンドリア外膜の透過性を亢進させる因子も幾つか報告されている。このように、アポトーシスシグナルをミトコンドリアに伝える機構には多様性があるようである。我々は、核、小胞体などからミトコンドリアに如何にしてアポトーシスシグナルが伝達されるのかに興味を持って解析を行っている。