|
|
|
The interleukin-6 cytokine family plays roles in a wide variety of tissues and organs, including the immune, hematopoietic, and nervous systems. Gp130 is a signal-transducing subunit shared by the receptors for the IL-6 family of cytokines. The binding of a ligand to its receptor induces the dimerization of gp130, leading to the activation of JAK tyrosine kinase and tyrosine phosphorylation of gp130. These events lead to the activation of multiple signal-transduction pathways, such as the STAT, Ras-MAPK, and PI-3 kinase pathways whose activation is controlled by distinct regions of gp130. We propose a model showing that the outcome of the signal transduction depends on the balance or interplay among the contradictory signal transduction pathways that are simultaneously generated through a cytokine receptor in a given target cell.
Since the molecular cloning of interleukin-6 (IL-6) in 1986 (1-6) , members of the IL-6 family of cytokines (7, 8) , including leukemia inhibitory factor (LIF) (9) , ciliary neurotrophic factor (CNTF) (10) , oncostatin M (OSM) (11) , interleukin-11 (IL-11) (12) and cardiotrophin-1 (CT-1) (13) have been molecularly cloned. During the last decade, many findings have been made concerning the structure and function of IL-6 family cytokines and their receptors (14). These findings, together with a large number of studies on many cytokines, have greatly contributed to the establishment of a variety of concepts about cytokines in general: the establishment of pleiotropy and redundancy as properties of cytokine function, the cytokine receptor super family, the sharing of a signal-transducing receptor subunit among several cytokine receptors, and the agonistic activity of certain soluble cytokine receptors. In fact, the IL-6 cytokine family plays pivotal roles in the immune, hematopoietic, nervous, cardiovascular, and endocrine systems, as well as in bone metabolism, inflammation, and acute phase response (5-8, 14-23) . Furthermore, they often exert overlapping biological activities; the molecular mechanism of this functional redundancy is explained at least in part by the sharing of gp130, a signal-transducing receptor subunit, among the receptors for the IL-6 cytokine family (7, 18, 24) . Important questions yet to be resolved are how a single cytokine can exert functional pleiotropy and how it can induce only a specific biological activity in a given target cell. For example, IL-6 induces macrophage differentiation and growth arrest in myeloid leukemic M1 cells whereas it induces growth in myeloma cells. Furthermore, the molecular mechanism by which each cytokine can exert a unique biological activity is still uncertain. Happily, studies on the signal-transduction mechanisms of interferons and erythropoietin have led to insight into the molecular mechanisms of signal transduction through cytokine receptors. It is now known that JAKs (Janus kinases) and STATs (signal transducer and activator of transcription) seem to play essential roles in cytokine function (25-28) . However, it is also true that the JAK-STAT pathway alone cannot explain all the events induced through cytokine receptors, and it is clear that other pathways involving src-family tyrosine kinases, RAS, mitogen-activated protein kinases (MAPK), phosphatidylinositol 3-kinase (PI-3 kinase), and as yet unidentified components participate, and that the interplay among them is critically involved in the biological activities of cytokines.
In this review, we describe recent progress in understanding signal transduction through gp130, with emphasis on the molecular mechanisms involved in the regulation of cell growth and differentiation.
Gp130 , a common signal transducing subunit of the IL-6 cytokine family receptors
IL-6, LIF, CNTF, OSM, IL-11, and CT-1 constitute the IL-6 cytokine family because of their functional redundancy, structural similarity, and sharing of a receptor subunit, gp130 (7, 8, 14, 17, 18, 23, 24, 29) . The IL-6 receptor (R) complex consists of an 80 kDa IL-6 binding molecule termed IL-6Ra, and a signal transducer, gp130, both belonging to the type I cytokine receptor super family (Figure 1) (30-33) .
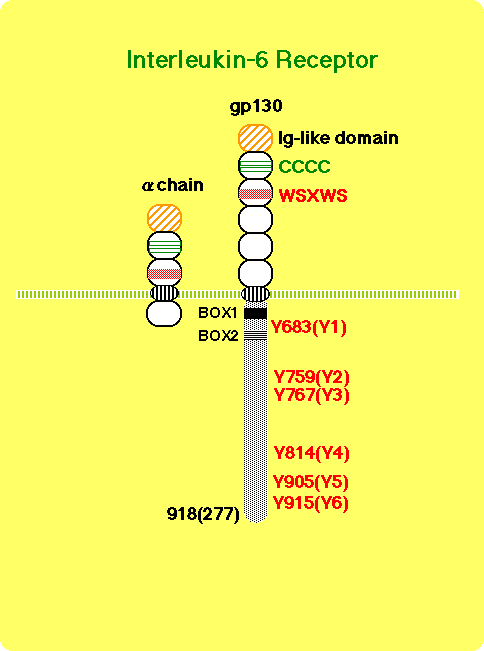
The cytoplasmic domain of IL-6Ra is not necessary for signal transduction (32, 34) . The binding of IL-6 to its receptor triggers the association of IL-6Ra with gp130 (31) , leading to the formation of a hexamer composed of two each of IL-6, IL-6Ra, and gp130 (35, 36) . Most importantly, it has been found that gp130 is shared among the receptors for CNTF, LIF, OSM, IL-11, and CT-1, and that it is critically involved in transducing signal transduction through these receptors ( Figure 2) (7, 14, 17, 18, 23, 24, 29) .
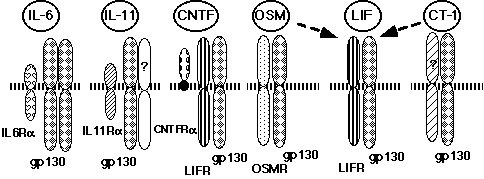
The sharing of a receptor subunit among different cytokine receptors is not a unique feature for the IL-6 cytokine family receptors, but rather a general feature of the cytokine receptor system. Granulocyte-macrophage colony stimulating factor (GM-CSF), IL-3, and IL-5 receptors share a common b subunit (37) . The b chain of the IL-2 receptor is shared by the IL-15 receptor, and the g chain of the IL-2 receptor (gc) is shared by the IL-4, IL-7, IL-9, and IL-15 receptors (38, 39) . Thus, the molecular mechanisms of redundancy in cytokine activity could be explained at least in part by the sharing of receptor subunits among several cytokine receptors. Interestingly, a complex of IL-6 and a soluble form of IL-6Ra can activate signal transduction in cells expressing only the gp130 receptor subunit. This type of arrangement is not unique to the IL-6 receptor system. IL-12 consists of a disulfide heterodimer of 40kD (p40) and 35kD (p35) subunits (40) . The peptide sequences of p35 and p40 resemble IL-6 and the soluble form of its receptor, respectively (41) , suggesting that IL-12 acts on target cells in a manner similar to the complex of IL-6 and soluble IL-6R. In fact, IL-12R is a member of the cytokine receptor super family, most closely related to gp130 (42) . Another example is a CNTFRa that is anchored to the cell membrane by a glycosyl-phosphatidyl inositol (GPI) linkage. The complex of soluble CNTFRa and CNTF acts on cells expressing LIFRb and gp130 (43) . Potential physiological roles for the soluble forms of CNTFRa are suggested by its presence in cerebrospinal fluid and its release from skeletal muscle in response to peripheral nerve injury. Furthermore, the complex of IL-11 and the soluble form of IL-11R can function through gp130 (44, 45) . Based on these facts, we originally proposed a novel mechanism by which the cytokine system generates functional diversity (Figure 3) (7, 46) .
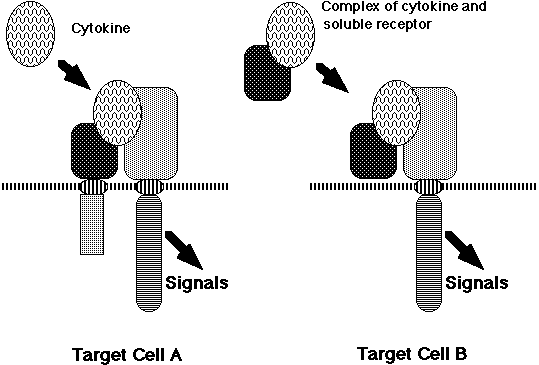
A complex consisting of a soluble cytokine receptor and its corresponding cytokine ligand acquires a different target specificity from the original cytokine and, therefore, it should express functions distinct from those of the original cytokine. Actually, double transgenic mice expressing human IL-6 and IL-6Ra showed myocardial hypertrophy (47) and extramedullary expansion of hematopoietic progenitor cells (48) , indicating that the complex of IL-6 and the soluble form of IL-6Ra acts on heart muscle cells and hematopoietic stem cells that express gp130, on which IL-6 alone cannot act. Thus, by forming a complex, IL-6 apparently acquired novel biological activities. This model may be also applied to the receptors for glial-cell-line-derived neurotrophic factor (GDNF) and neurturin (NTN). Both the GDNF and NTN receptors consist of a ligand-specific GPI-anchored a-chain and a common signal-transducing receptor subunit, Ret, which is a receptor tyrosine kinase (49-52) . This novel mechanism may be applied to a wide range of receptor systems. This mechanism contributes to generating the functional diversity of cytokines and may also play pathological roles in various diseases, since an increase in the serum-soluble form of various cytokine receptors has been reported to occur in a variety of diseases. Furthermore, novel drugs could be designed based on this model. In an elegant experiment, Rose-John and his colleagues developed a bioactive designer cytokine composed of soluble IL-6Ra and IL-6 linked each other by a flexible peptide chain (53) . The designer cytokine can effectively expand hematopoietic progenitor cells in the presence of SCF and IL-3.
Signal transduction through gp130: from membrane to the nucleus.
One of the hottest issues until 1993 was the identification of catalytic molecules that associate with cytokine receptors. This issue was resolved by the findings that Tyk2, a JAK family tyrosine kinase, is critical for IFNa signaling (54) , and that JAK2 is associated with the erythropoietin and growth-hormone receptors, and is activated by these ligands (55, 56) . Since then, several JAK family tyrosine kinases (JAK1, JAK2, JAK3, Tyk2) have been shown to be involved in the signal transduction of cytokines and hormones (25, 28) . Cytokines induce receptor aggregation, resulting in the activation of JAK family tyrosine kinases. These events eventually induce the tyrosine phosphorylation of STAT, which was originally identified as an interferon-activated transcription factor by Darnell and his colleagues (26, 27) . Phosphorylated STATs are dimerized and translocated into the nucleus, leading to the expression of genes with STAT recognition sites.
JAK1, JAK2, and Tyk2 associate constitutively with gp130 and are tyrosine-phosphorylated in response to IL-6, CNTF, LIF, OSM, or IL-11 (57-60) . JAK1 is considered to be a major kinase among this family to activate STAT3 through gp130 (61) . IL-6 activates STAT3, STAT1, and STAT5 (62-67) . Two types of IL-6 responsive elements (REs) have been identified in the genes encoding acute-phase proteins: type I IL-6 RE, which is a binding site for NF-IL6 and other members of the C/EBP family (68, 69) and type II IL-6 RE. IL-6 triggers the rapid activation of a nuclear factor, termed acute-phase response factor (APRF) binding to type II IL-6 RE (70) . The purification and molecular cloning of APRF revealed that APRF has marked similarity with STAT1 (62) and is identical with STAT3 (63) . In parallel with these studies, Nakajima et al. (71) identified an IL-6 responsive element in the junB gene promoter (JRE-IL6). This element consists of a putative Ets-binding site (JEBS) and a cAMP-responsive element (CRE)-like site. Both sites in JRE-IL6 are essential for the IL-6-dependent junB expression. Furthermore, the IL-6-inducible JEBS-binding protein complex mainly contains STAT3, although the JEBS is a low-affinity binding site for STAT3 as compared with APRE (64, 65) . IL-6 induces the formation of a complex consisting of STAT3 and a CRE-like site binding protein with a molecular weight of 36kDa on the JRE-IL6 (72) . The cooperation between the STAT3-binding site and the CRE-like site is also observed in both the IL-6/IFNg responsive element in the IRF-1 promoter (72) and the IL-6 responsive element of the stat3 gene (Ichiba et al, manuscrupt submitted). The formation of such a binding complex seems to be important for STAT to act on a low-affinity binding site, such as the JEBS, and may contribute to generating the diversity of STAT protein target genes. In addition to the tyrosine phosphorylation of STAT3 by JAKs, the H7-sensitive pathway, most likely a serine/threonine kinase, is required for the transactivation through the JRE-IL6 and APRE (65, 71) , although H7 does not inhibit the IL-6-induced appearance of the DNA binding complex on the APRE or JEBS (65) . In certain cell lines, it has been shown that STAT3 requires phosphorylation on serine to form a STAT3-STAT3 homodimer when bound to a low-affinity STAT3 binding site (73) , further suggesting the involvement of a serine/threonine kinase in the STAT signal pathway. In fact, maximal activation of transcription by STAT1 and STAT3 requires both tyrosine and serine phosphorylation (74) . Furthermore, it has been suggested that ERK2 MAPK is involved in IFN-b-induced activation of STAT1: treatment of cells with IFN-b induces the activation of ERK2 and the association of ERK2 with STAT1. In addition, expression of a dominant negative ERK2 inhibites IFN-b-induced transcription (75) . However, it is still controversial whether ERK MAPKs are actually involved in the activation of STAT. It is possible that as yet unidentified members of the MAPK family are responsible for the activation of STAT molecules. The p85 regulatory subunit of PI-3 kinase binds to tyrosine-phosphorylated STAT3 in response to IFN-a and subsequently undergoes tyrosine phosphorylation (76) . Because PI-3 kinase has endogenous serine kinase activity and activates a serine kinase cascade, and because a PI-3 kinase inhibitor, wortmannin, inhibits IFN-a-induced formation of a slowly migrating band of STAT3, which is phosphorylated on both serine and tyrosine, it has been suggested that PI-3 kinase may promote the serine phosphorylation of STAT3 (76) .
In response to cytokine stimulation, STATs enter the nucleus, where they activate transcription. However, the molecular mechanisms by which STAT proteins are transported into the nucleus remains unknown. STAT1 is actively transported through the nuclear pores in an IFN-g-dependent manner, and tyrosine (Tyr701) phosphorylation of STAT1 is required for its nuclear importation. Furthermore, a nuclear small GTP-binding protein Ran, which is an essential factor for active nuclear protein transport, is involved in, and its GTP hydrolysis activity is required for, the IFN-g-dependent nuclear transport of STAT1 (77) .
In addition to the JAK/STAT pathway, multiple signaling molecules are tyrosine phosphorylated in response to the IL-6 family of cytokines. CNTF, LIF, OSM, and IL-6 induce tyrosine phosphorylation of phospholipase C g, SHP-2 (a phosphotyrosine phosphatase, also called PTP1-D, SHPTP-2, PTP2C, and Syp) which is a mammalian homolog of Drosophila corkscrew (CWS), pp120, Shc, Grb2, Raf-1, and ERK1 and ERK2 (78) . IL-11 induces tyrosine phosphorylation of SHP-2 in mouse 3T3-L1 cells. Furthermore, SHP-2 is inducibly associated with gp130 (79, 80) and JAK2 (79) . The Ras-MAPK pathway is activated by the IL-6 cytokine family (78, 80-85) . The activation of the Ras-MAPK pathway is possibly mediated by SHP-2 (80, 84) and/or Shc (83, 86) , which bind a Grb2-SOS complex. Gab1 is also involved in MAPK activation as described below (Takahashi-Tezuka et al, manuscript submitted). Src-family tyrosine kinases, such as Btk, Tec, Fes, and Hck (86-89) are activated by the IL-6 cytokine family, as well as by a variety of other cytokines (39) . Among them, Tec and Btk associate with and are possibly activated by JAKs, and Tec may be one of the adaptor molecules linking the cytokine receptor to PI-3 kinase (90) .
Distinct cytoplasmic regions of gp130 are involved in different signal-transduction pathways.
Human gp130 has 277 amino acid residues in its cytoplasmic domain, which contains two motifs conserved among the cytokine receptor family, termed box 1 and box 2 (Fig. 1) (32, 91, 92) . The membrane-proximal region containing box1 and box2 is sufficient for JAK to be activated through gp130 (93) . Furthermore, the same membrane-proximal region, containing 68 amino acid residues, is sufficient for the induction of the c-myc gene (80) , activation of STAT5 (67) , and tyrosine phosphorylation of Gab1 (which has structural similarities to Drosophila DOS, daughter of sevenless) (Takahashi-Tezuka, manuscript submitted) (Fig. 4).
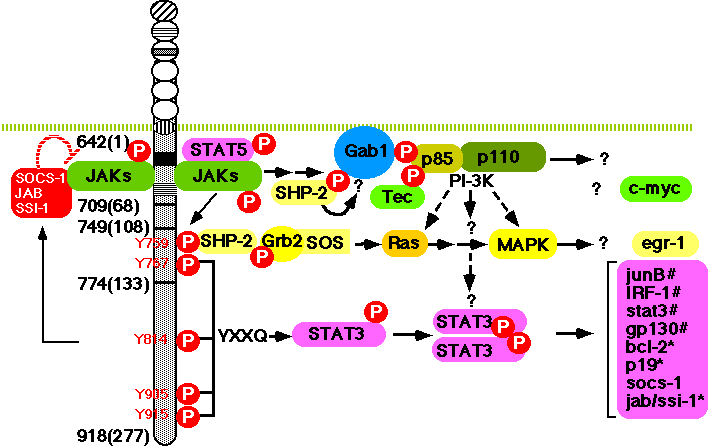
Gp130 has six tyrosine residues in its cytoplasmic domain but none of them is required for the above responses. Rather, the activation of STAT3 and the tyrosine phosphorylation of SHP-2 are dependent on tyrosine residues of gp130 (Fig. 4). Tyrosine 759, the second tyrosine (counting away from the cell membrane towards the cytoplasmic tail), is required for the tyrosine phosphorylation of SHP-2 and its binding to gp130 (80, 94) , the association between SHP-2 and Grb2 and the full activation of MAPK (80) . Gab1 is tyrosine phosphorylated in response to various cytokines, such as IL-6, IL-3, IFN-a, and IFN-g. Upon such stimulus, Gab1 forms a complex with PI-3 kinase and SHP-2. Overxpression of Gab1 enhances gp130-dependent ERK2 MAPK activation, and this activation can be inhibited by either mutating tyrosine 759, the SHP-2 binding site of gp130, or by expressing a dominant negative p85 subunit of PI-3 kinase. These observations suggest that Gab1 acts as an adaptor molecule linking gp130 to PI-3 kinase and MAPK (Takahashi-Tezuka and Yoshida et al, Mol Cell Biol. in press). Gab1 is also tyrosine phosphorylated in response to EGF, insulin, and c-Met stimulation; it has binding sites for PLC-g, PI-3 kinase, SHP-2, and Grb2 (95, 96) and shows structural similarities with IRS-1, IRS-2, and Drosophila DOS. These DOS-related family molecules may act as universal docking molecules linking a variety of receptors to downstream signaling molecules. In fact, IRS-1 is tyrosine phosphorylated in response to IL-2, IL-4, IL-7, IL-9, IL-15, Oncostatin M, and interferons in addition to insulin (97-100) . IRS-2 acts as an adaptor molecule linking growth hormone receptor to PI-3 kinase. Growth hormone induces the association of IRS-2 with the p85-kDa regulatory subunit of PI-3 kinase and with SHP-2 (101) . The IL-4-induced-tyrosine phosphorylation of IRS-1 is dependent on JAK (99) and the phosphorylation of tyrosine 497 of the IL-4 receptor a chain (97) , whereas the growth hormone-induced tyrosine phosphorylation of IRS-2 is independent of the phosphorylation of the receptor's tyrosine residues and the regions of the growth hormone receptor necessary for IRS-2 tyrosine phosphorylation are the same as those required for JAK2 association and tyrosine phosphorylation (101) . The observation is similar to the gp130-mediated tyrosine phosphorylation of Gab1 (Takahashi-Tezuka and Yoshida et al, Mol. Cell. Biol. in press). IRS-1 is found to be associated with JAK1 (100) . The results suggest that JAK may directly induce the tyrosine phosphorylation of IRS-1, IRS-2, and Gab1 (Fig. 4).
The activation of STAT3 is dependent on the phosphorylation of any one of the four tyrosines (Y767, Y814, Y905, Y915) in the carboxy terminus of gp130 that have a glutamine residue at the third position behind tyrosine (Y-X-X-Q) (Fig. 4) (94, 102) . In accordance with the fact that STAT3 is involved in the activation of type II acute-phase genes, the membrane-proximal region, containing 133 amino acids including Y767, is the minimal region of gp130 necessary for the activation of IL-6 responsive acute-phase genes (103) (Fig. 5). As shown in Fig. 4, multiple signal transduction pathways could be generated through distinct cytoplasmic regions of gp130.
Signal transduction pathways regulating cell growth, differentiation and death
IL-6 is a typical pleiotropic cytokine. For example, IL-6 induces the differentiation of B cells to antibody-producing plasma cells (1) , T-cell growth and differentiation (in particular, Th2 development (104) ), the differentiation of myeloid leukemic cell lines into macrophages (105-107) , and the neural differentiation of PC12 cells (108) . IL-6 is a growth factor for myeloma/plasmacytoma (109-112) . The questions to be resolved include how a single cytokine can exert functional pleiotropy and how it can induce only a specific biological activity in a given target cell.
Yamanaka et al. (102) showed that the membrane-proximal region of gp130, containing 133 amino acids, is sufficient to generate the signals for growth arrest, macrophage differentiation, down-regulation of c-myc and c-myb, induction of junB,IRF1, and egr-1, and the activation of STAT3. The region between 108 and 133 amino acids contains two tyrosine residues (Fig. 4 and Fig. 5): one at amino acid position 767(126 residues from the membrane) with the YXXQ motif, and another without the motif at amino acid position 759 (118 residues from the membrane). Tyrosine 759 is essential for gp130-mediated egr-1 gene induction. Tyrosine 767, which has the motif, has been shown to be essential for truncated gp130 (containing the membrane-proximal 133 amino acids) to generate the signals, not only for STAT3 activation, but also for growth arrest and differentiation, accompanied by the down-regulation of c-myc and c-myb and the immediate early induction of junB and IRF1 (102) . These results suggest that STAT3 plays an essential role in the signals for growth arrest and differentiation. This possibility was confirmed by Nakajima et al. (113) , who showed that dominant-negative forms of STAT3 inhibite both IL-6-induced growth arrest and macrophage differentiation in the M1 transformants. Blocking STAT3 activation inhibites IL-6-induced repression of c-myb and c-myc, but not IL-6-induced egr-1 expression. The involvement of STAT3 in the IL-6-induced macrophage differentiation of M1 cells (114) , and G-CSF-induced neutrophil differentiation (115) has also been demonstrated. Furthermore, IL-6 induces p19INK4D and its induction is suppressed by dominant-negative forms of Stat3 (Narimatsu et al, BBRC, 238:764-768, 1997). Therefore, this is another example that STAT is involved in cell growth arrest by inducing CDK inhibitors as previously shown in the case of IFNg-induced p21WAF1/CIP1 (116) . Thus both Stat3-regulated events: the repression of c-myc and c-myb and the induction of p19INK4D, are likely to be involved in IL-6-induced growth arrest in M1 cells. The possible involvement of CDK inhibitors, p18INK4c and p21WAF1/CIP1 in the IL-6-induced differentiation of B cells to plasma cells has also been suggested (117) . Furthermore, IL-6 enhances the growth of M1 cells when STAT3 is suppressed. Thus, IL-6 generates growth-enhancing signals, and growth arrest- and differentiation-inducing signals at the same time, but the former are only apparent when STAT3 activation is suppressed. Therefore, the outcome of the signal transduction depends on the balance or interplay among the contradictory signal-transduction pathways that are simultaneously activated through gp130 in M1 cells as discussed later.
For the growth signal, it was shown that a 65-amino acid region of gp130 proximal to the transmembrane domain is sufficient for transient 3H-Tdr uptake, by using gp130-transfectants of an IL-3-dependent Pro-B-cell line BAF/B03 (18, 92) . However, Fukada et al. (80) showed that a membrane-proximal region of 68 amino acids is not sufficient to induce 3H-Tdr uptake when cells are starved of IL-3, although this region is sufficient to induce c-myc gene activation. The membrane-proximal region containing 133 amino acid residues is required and sufficient for cell growth (Fig. 5).
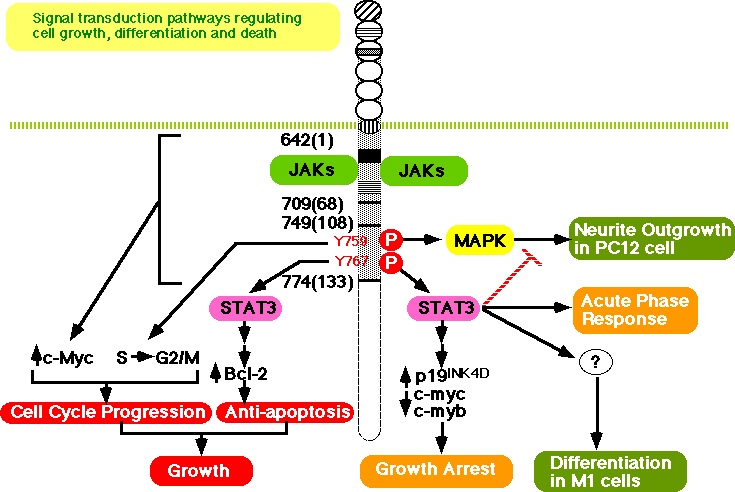
Furthermore, at least two distinct signals are required for gp130-induced cell growth: one is a cell-cycle progression signal dependent on tyrosine 759, the SHP-2 binding site required for MAPK activation, and the other is an anti-apoptotic signal dependent on tyrosine 767. The latter signal is mediated by STAT3. Tyrosine 759 is essential for the transition from S to G2/M but dispensable for the anti-apoptotic signal. Since mutation of tyrosine 759 severely abrogates the gp130-induced activation of MAPK, the results suggest the involvement of MAPK in cell-cycle progression. This is consistent with the observation that the expression of active form of N-ras in IL-6-dependent myeloma cells results in significant IL-6-independent growth, as well as augmentation of growth at suboptimal concentrations of IL-6 (118) , and v-Ha-ras partially alleviates the requirement of embryonic stem (ES) cells for LIF to proliferate (86) . On the other hand, mutations of tyrosine 767 in the context of the membrane-proximal 133 amino acid residues of gp130 disrupt STAT3 activation and result in apoptosis and the loss of bcl-2 induction. Moreover, dominant-negative STAT3 inhibits both gp130-mediated anti-apoptotic signals and bcl-2 induction (80) . Consistent with this, high-level expression of an exogenous bcl-2 gene, introduced into IL-6-dependent B9 myeloma cells, is able to suppress apoptotic death following cytokine deprivation, although this particular myeloma cell line expresses cellular bcl-x mRNA but not bcl-2 mRNA in response to IL-6 (119) . The results indicate that STAT3 is involved in the generation of an anti-apoptotic signal, most likely through the induction of the bcl-2 or bcl-x gene.
IL-6 has been reported to induce neurite outgrowth in PC12 cells (108) . However, this effect is limited to certain lines of PC12 cells, and most PC12 cell lines do not differentiate after IL-6 stimulation (120, 121) . Ihara et al. (121) found that IL-6 could induce the differentiation of PC12 cells if the cells were pre-treated with NGF for one hour. Consistent with this, IL-6 requires stimulation with low concentrations of NGF to induce neurite outgrowth in PC12 cells (122) . Ihara and her colleagues further elucidated the molecular mechanisms by which IL-6 induces neurite outgrowth in PC12 cells pretreated with NGF, and showed that the membrane-proximal region of gp130, containing 133 amino acid residues, is the minimal region required for the neurite outgrowth. Furthermore, they showed that tyrosine 759, the binding site for SHP-2 and required for MAPK activation, is essential for neurite outgrowth, but tyrosine 767, the binding site for STAT3, is inhibitory. Actually, stimulation through a variant of the truncated gp130 that bears a mutation at tyrosine 767 induces neurite outgrowth without pretreatment of NGF. Furthermore, in the presence of dominant negative STAT3, IL-6 alone can induce neurite outgrowth in PC12 cells, indicating that STAT3 negatively regulates neurite outgrowth in PC12 cells (123) (Fig. 5). The negative effect of STAT3 on cell differentiation is also implicated in LIF activity: STAT3 is critical for the LIF-mediated signaling pathway that maintains pluripotent proliferation of embryonic stem cells (124) .
How can a single cytokine exert a variety of functions and how can a given target cell respond specifically ?
Cytokines exert a variety of biological activities through specific receptors. Since the expression patterns of each cytokine receptor and of the cytokine itself are different, and since each cytokine receptor has different binding affinities for a variety of signaling molecules, such as STAT proteins which are key molecules responsible for the expression of the specificity of cytokine signal-transduction pathway as previously discussed (14, 27, 94, 125, 126) , each cytokine is capable of expressing a unique biological activity, although functional redundancy is observed among related cytokines, such as the IL-6 cytokine family. On the other hand, it is still difficult to explain precisely how a single cytokine, for example IL-6, can exert distinct biological activities on different target cells. However, one may consider several points. First, different sets of signal-transduction pathways (despite the existence of partial redundancy among them), could simply be acitvated in different targets through a given cytokine receptor such as gp130, due to differences in the expression pattern of each signaling molecule (Fig. 6A).
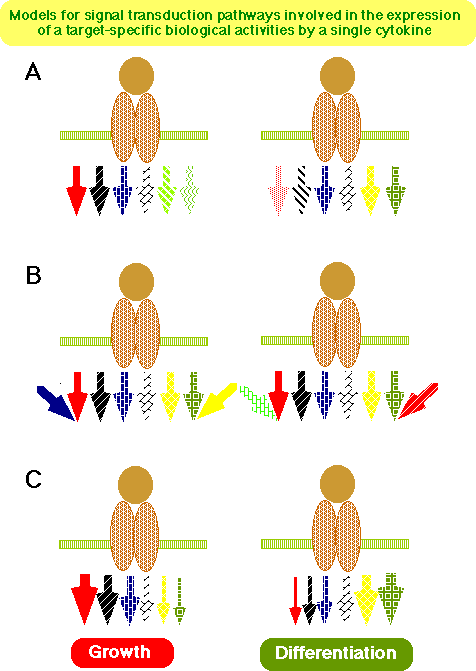
Second, even if gp130 induces the same set of signal transduction pathways in different targets, each target cell could respond to gp130 stimulation differently because the expression and or activation state of other molecules affecting each signal transduction pathway negatively or synergistically is different or because the final transcriptional activation of target genes of the signal transduction pathway is different among different targets (Fig. 6B). Third, the balance or interplay (inhibitory or synergistic interaction) among the signaling pathways could determine the eventual outcome of the signal transduction through the receptor in a given target cell (Fig. 6C). Relevant findings obtained through the studies on the gp130-mediated signals are: 1) gp130-stimulation can simultaneously induce opposite signals-- i.e., growth enhancing and growth suppressing signals are induced in M1 cells as described above-- (113) and 2) gp130 can deliver at the same time both positve and negative signals affecting neurite outgrowth in PC12 cells (123) . These findings seem to be quite surprising since one might reasonably expect that a cytokine exerts distinct biological activities in different target cells simply because the signal transduction pathways activated by the cytokine are different or because the state of differentiation of each target is different (Fig. 6 A and B), and therefore the cytokine would only induce a growth signal in growth-responding cells, while it induced a growth suppressing signals in cells that stop growing in response to the cytokine stimulation. A similar observation is reported in TNF stimulation, which elicits simultaneously both apoptotic signals through the caspase cascade and anti-apoptotic signals through NF-kB activation (127) . In short, the cytokine receptor can simultaneously induce contradictory intracellular signaling pathways, at least in certain cells, and the balance and or interplay of each pathway could determine the final outcome of the stimulation (Fig. 6C). Acutually, it is likely that the variable combinations of these mechanisms are involved in the determination of the final outcome of the cytokine signaling.
Such events could be influenced by a variety of factors that determine the duration and intensity of each signal generated by all the activated pathways. In this sense, the presence of up- or down-regulating mechanisms involved in each signaling pathway may be crucial to determining the balance. The activation state of STAT3 in IL-6-stimulated M1 cells persists for as long as 24 hours after the stimulation (113) . Such a persistent activation of STAT3 is not observed in other cells, such as a pro-B cell line, BAF/B03. Similarly there is sustained activation of MAPK in NGF-stimulated PC12 cells that is not observed in PC12 cells stimulated by EGF, which cannot induce neurite outgrowth (128, 129) . Such prolonged activation of a particular signal transduction pathway should affect the outcome of the signal transduction. The sustained activation of STAT3 in M1 cells may be induced by either the absence of a negative regulator of STAT3, such as a postulated STAT phosphotyrosine phosphatase (130) , or the upregulation of STAT3. In fact, the Stat3 gene is autoregulated by STAT3 in M1 cells (Ichiba et al, manuscript submitted) and this may partly contribute to the sustained activation of STAT3 in M1 cells. Concerning a negative regulator for STAT, the natural existence of potentially dominantly suppressive variants of STAT3 and STAT5 has been reported (131, 132) . Since NGF-pretreatment inhibits the gp130-mediated tyrosine phosphorylation of STAT3 in PC12 cells, NGF may induce a STAT3 inhibitor, including a phosphotyrosine phophatase (123) . Phosphotyrosine phosphatases are critical negative or positive regulators for cytokine and growth factor-mediated signal transduction pathway. SHP-1, an SH2 domain containing a phosphotyrosine phosphatase, is thought to act as a negative regulator for erythropoietin receptor-mediated signal transduction by inactivating Jak2 (133) , whereas SHP-2 is thought to act as a positive regulator (134) . In addition to these, the family molecules that can bind to SHP-2, SHP-1, and Grb2 have been cloned: these are the signal-regulatory protein (SIRP) family (135) and SHP substrate 1 (SHPS-1) (136) which is a member of the SIRPa family. SIRPa1 is a substrate for activated receptor tyrosine kinases and its tyrosine-phosphorylated form binds SHP-2 through SH2 interactions and acts as its substrate. It has negative regulatory effects on insulin, EGF, and PDGF-induced growth, most likely through the inhibition of MAPK activity (135) . Furthermore, it has been reported recently that STAT3 induces a SH2-domain-containing molecule designated as SOCS-1/JAB/SSI-1, which is structurally related to CIS, a cytokine-inducible SH2 protein (137-139) . SOCS-1/JAB/SSI-1 can bind JAK and inhibit its kinase activity and thereby suppress the tyrosine phosphorylation of gp130 and subsequent activation of STAT (Fig. 4).
The identification of novel molecules involved in signal-transduction pathways, elucidation of the molecular mechanisms of each signal transduction pathway and the interplay among them, and the clarification of both in vitro and in vivo roles of each signaling molecules would clarify our concept of how signal transduction pathways are involved in the regulation of cell growth, differentiation, and survival.
Acknowledgments
We thank many colleagues who contributed to our recent studies described in this review. This study was supported in part by a Grant-in-Aid for COE Research of the Ministry of Education, Science, Sports and Culture in Japan.
ü@
1. Hirano T, Yasukawa K, Harada H, et al. Complementary DNA for a novel human interleukin (BSF-2) that induces B lymphocytes to produce immunoglobulin. Nature 1986;324:73-76.
2. Zilberstein A, Ruggieri R, Korn JH, Revel M. Structure and expression of cDNA and genes for human interferon-beta-2, a distinct species inducible by growth-stimulatory cytokines. EMBO J. 1986; 5:2529-2537.
3. Haegeman G, Content J, Volckaert G, Derynck R, Tavernier J, Fiers W. Structural analysis of the sequence encoding for an inducible 26-kDa protein in human fibroblasts. Eur J Biochem 1986;159:625-632.
4. May LT, Helfgott DC, Sehgal PB. Anti-b-interferon antibodies inhibit the increased expression of HLA-B7 mRNA in tumor necrosis factor-treated human fibroblasts: Structural studies of the b2 interferon involved. Proc Natl Acad Sci USA 1986; 83:8957-8961.
5. Sehgal PB, Grienger G, Tosato G, eds. Regulation of the Acute Phase and Immune Responses: Interleukin-6. 1989. Ann New York Acad Sci; vol 557.
6. Hirano T, Kishimoto T. Interleukin 6. In: Sporn MB, Roberts AB, eds. Handbook of Experimental Pharmacology. Peptide growth factors and their receptors. Berlin: Springer-Verlag, 1990:633-665. vol 95/1.
7. Hirano T, Matsuda T, Nakajima K. Signal transduction through gp130 that is shared among the receptors for the interleukin 6 related cytokine subfamily. Stem Cells 1994;12:262-277.
8. Mackiewicz A, Koj A, Sehgal PB, eds. Interleukin-6-type cytokines. New York: 1995. Ann New York Acad Sci; vol 762.
9. Gearing D, Gough NM, King JA, et al. Molecular cloning and expression of cDNA encoding a murine myeloid leukaemia inhibitory factor (LIF). EMBO J. 1987;6:3995-4002.
10. Lin L-FH, Mismer D, Lile JD, et al. Purification, cloning, and expression of ciliary neurotrophic factor (CNTF). Science 1989;246:1023-1025.
11. Malik N, Kalestad JC, Gunderson NL, et al. Molecular cloning, sequence analysis, and functional expression of a novel growth regulator, oncostatin M. Mol. Cell. Biol. 1989;9:2847-2853.
12. Paul SR, Bennett F, Calvetti JA, et al. Molecular cloning of a cDNA encoding interleukin 11, a stromal cell-derived lymphopoietic and hematopoietic cytokine. Proc. Natl. Acd. Sci. USA 1990;87:7512-7516.
13. Pennica D, King KL, Shaw KL, et al. Expression cloning of cardiotrophin-1, a cytokine that induces cardiac myocyte hypertrophy. Proc. Natl. Acad. Sci. USA 1995;92:1142-1146.
14. Hirano T. Interleukin 6 and its receptor: Ten years later. Intern Rev Immunol. 16:249-284, 1998(Full Text)(PubMed)
15. Metcalf D. The leukemia inhibitory factor (LIF). Int. J. Cell Cloning 1991;9:95-108.
16. Patterson PH, Nawa H. Neuronal differentiation factors/cytokines and synaptic plasticity. Cell 1993;72:123-137.
17. Pennica D, Wood WI, Chien KR. Cardiotrophin-1: a multifunctional cytokine that signals via LIF receptor-gp 130 dependent pathways. Cytokine Growth Factor Rev 1996;7:81-91.
18. Kishimoto T, Akira S, Narazaki M, Taga T. Interleukin-6 family of cytokines and gp130. Blood 1995;86:1243-1254.
19. Hirano T. The biology of interleukin-6. Chem Immunol 1992;51:153-80.
20. Hilton DJ. LIF: lots of interesting functions. Trends-Biochem-Sci 1992;17:72-76.
21. Yang YC. Interleukin 11: an overview. Stem Cells 1993;11:474-486.
22. Ip NY, Yancopoulos GD. The neurotrophins and CNTF: two families of collaborative neurotrophic factors. Annu Rev Neurosci 1996;19:491-515.
23. Gearing DP. The leukemia inhibitory factor and its receptor. Adv Immunol 1993;53:31-58.
24. Hibi M, Nakajima K, Hirano T. IL-6 cytokine family and signal transduction: a model of the cytokine system. J. Mol. Med. 1996;74:1-12.
25. Ihle JN, Witthuhn BA, Quelle FW, et al. Signaling by the cytokine receptor superfamily: JAKs and STATs. Trends Biochem Sci 1994;19:222-227.
26. Darnell JJ, Kerr IM, Stark GR. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994;264:1415-21.
27. Schindler C, Darnell JE, Jr. Transcriptional responses to polypeptide ligands:The JAK-STAT pathway. Annu. Rev. Biochem. 1995;64:621-651.
28. Ihle JN, Witthuhn BA, Quelle FW, Yamamoto K, Silvennoinen O. Signaling through the hematopoietic cytokine receptors. Annu Rev Immunol 1995;13:369-398.
29. Davis S, Yancopoulos GD. The molecular biology of the CNTF receptor. Curr-Opin-Cell-Biol 1993;5:281-285.
30. Yamasaki K, Taga T, Hirata Y, et al. Cloning and expression of the human interleukin-6 (BSF-2/IFN beta 2) receptor. Science 1988;241:825-828.
31. Taga T, Hibi M, Hirata Y, et al. Interleukin-6 triggers the association of its receptor with a possible signal transducer, gp130. Cell 1989;58:573-581.
32. Hibi M, Murakami M, Saito M, Hirano T, Taga T, Kishimoto T. Molecular cloning and expression of an IL-6 signal transducer, gp130. Cell 1990;63:1149-1157.
33. Bazan JF. Haemopoietic receptors and helical cytokines. Immunol Today 1990;11:350-354.
34. Sugita T, Totsuka T, Saito M, et al. Functional murine interleukin 6 receptor with the intracisternal A particle gene product at its cytoplasmic domain. Its possible role in plasmacytomagenesis. J. Exp. Med 1990;171(6):2001-2009.
35. Ward LD, Howlett GJ, Discolo G, et al. High affinity interleukin-6 receptor is a hexameric complex consisting of two molecules each of interleukin-6, interleukin-6 receptor, and gp-130. J. Biol. Chem. 1994;269:23286-23289.
36. Paonessa G, Graziani R, Serio AD, et al. Two distinct and independent sites on IL-6 trigger gp130 dimer formation and signalling. EMBO J. 1995;14:1942-1951.
37. Miyajima A, Kitamura T, Harada N, Yokota T, Arai K. Cytokine receptors and signal transduction. Annu. Rev. Immunol. 1992;10:295-339.
38. Sugamura K, Asao H, Kondo M, et al. The common gamma-chain for multiple cytokine receptors. Adv Immunol 1995;59:225-227.
39. Taniguchi T. Cytokine signaling through nonreceptor protein tyrosine kinase. Science 1995;268:251-255.
40. Kobayashi M, Fitz L, Ryan M, et al. Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J. Exp. Med. 1989;170:827-845.
41. Gearing DP, Cosman D. Homology of the p40 subuniot of natural killer cell stimulatory factor (NKSF) with the extracellular domain of the interleukin-6 receptor. Cell 1991;66:9-10.
42. Chua AO, Chizzonite R, Desai BB, et al. Expression cloning of a human IL-12 receptor component. A new member of the cytokine receptor superfamily with strong homology to gp120. J. Immunol. 1994;153:128-136.
43. Davis S, Aldrich TH, Ip NY, et al. Released form of CNTF receptor a components as a soluble mediator of CNTF responses. Science 1993;259:1736-1739.
44. Baumann H, Wang Y, Morella KK, et al. Complex of the soluble IL-11 receptor and IL-11 acts as IL-6-type cytokine in hepatic and nonhepatic cells. J. Immunol 1996;157:284-290.
45. Neddermann P, Graziani R, Ciliberto G, Paonessa G. Functional expression of soluble human interleukin-11 (IL-11) receptor alpha and stoichiometry of in vitro IL-11 receptor complexes with gp130. J Biol Chem 1996;271:30986-30991.
46. Hirano T. Interleukin 6. In: Thomson AW, ed. The Cytokine Handbook, The second edition. London: Academic Press, 1994:145-168.
47. Hirota H, Yoshida K, Kishimoto T, Taga T. Continuous activation of gp130, a signal transducing receptor component for interleukin 6-related cytokines, causes myocardial hypertrophy in mice. Proc. Natl. Acad. Sci. USA. 1995;92:4862-4866.
48. Peters M, Schirmacher P, Goldschmitt J, et al. Extramedullary expansion of hematopoietic progenitor cells in interleukin (IL)-6-sIL-6R double transgenic mice. J Exp Med 1997;185:755-766.
49. Treanor JJS, Goodman L, de Sauvage F, et al. Characterization of a multicomponent receptor for GDNF. Nature 1996;382:80-83.
50. Jing S, Wen D, Yu Y, et al. GDNF-Induced Activation of the Ret Protein Tyrosine Kinase Is Mediated by GDNFR-a, a Novel Receptor for GDNF. Cell 1996; 85:1113-1124.
51. Klein RD, Sherman D, Ho W-H, et al. A GPI-linked protein that interacts with Ret form a candidate neurturin receptor. Nature 1997;387:717-721.
52. Buj-Bello A, Adu J, Pinon LGP, et al. Neurturin responsiveness requires a GPI-linked receptor and the Ret receptor tyrosine kinase. Nature 1997;387:721-724.
53. Fisher M, Goldschmitt J, Peschel C, et al. A bioactive designer cytokine for human hematopoietic progenitor cell expansion. Nature Biotechnology 1997;15:142-145.
54. Velazquez L, Fellous M, Stark GR, Pellegrini S. A protein tyrosine kinase in the interferon a/b signaling pathway. Cell 1992;70:313-322.
55. Witthuhn BA, Quelle FW, Silvennoinen O, et al. Jak2 associates with the erythropoietin receptor and is tyrosine phosphorylated and activated following stimulation with erythropoietin. Cell 1993;74:227-236.
56. Argetsinger LS, Campbell GS, Yang X, et al. Identification of Jak2 as a growth hormone receptor-associated tyrosine kinase. Cell 1993;74:237-244.
57. Lutticken C, Wegenka UM, Yuan J, et al. Association of transcription factor APRF and protein kinase Jak1 with the interleukin-6 signal transducer gp130. Science 1994;263:89-92.
58. Stahl N, Boulton TG, Farruggella T, et al. Association and activation of Jak-Tyk kinases by CNTF-LIF-OSM-IL-6 beta receptor components. Science 1994;263:92-5.
59. Matsuda T, Yamanaka Y, Hirano T. Interleukin-6-induced tyrosine phosphorylation of multiple proteins in murine hematopoietic lineage cells. Biochem. Biophys. Res. Commun 1994;200:821-828.
60. Berger LC, Hawley TS, Lust JA, Goldman SJ, Hawley RG. Tyrosine phosphorylation of JAK-TYK kinases in malignant plasma cell lines growth-stimulated by interleukins 6 and 11. Biochem. Biophys. Res. Commun 1994;202:596-605.
61. Guschin D, Rogers N, Briscoe J, et al. A major role for the protein kinase JAK1 in the JAK/STAT signal transduction pathway in response to interleukin-6. EMBO J. 1995;14:1421-1429.
62. Akira S, Nishio Y, Inoue M, et al. Molecular cloning of APRF, a novel IFN-stimulated gene factor 3 p91- related transcription factor involved in the gp130-mediated signaling pathway. Cell 1994;77:63-71.
63. Zhong Z, Wen Z, Darnell JE. Stat3: a STAT family member activated by tyrosine phosphorylation in response to epidermal growth factor and interleukin-6. Science 1994;264:95-98.
64. Fujitani Y, Nakajima K, Kojima H, Nakae K, Takeda T, Hirano T. Transcriptional activation of the IL-6 response element in the junB promoter is mediated by multiple Stat family proteins. Biochem. Biophys. Res. Commun 1994;202:1181-1187.
65. Nakajima K, Matsuda T, Fujitani Y, et al. Signal transduction through IL-6 receptor: involvement of multiple protein kinases, stat factors, and a novel H7-sensitive pathway. Ann N Y Acad Sci 1995;762:55-70.
66. Lai CF, J. Ripperger, K. K. Morella, et al. STAT3 and STAT5B are targets of two different signal pathways activated by hematopoietin receptors and control transcription via separate cytokine response elements. J. Biol. Chem. 1995; 270:23254-23257.
67. Fujitani Y, Hibi M, Fukada T, et al. An alternative pathway for STAT activation that is mediated by the direct interaction between JAK and STAT. Oncogene 1997;14:751-761.
68. Akira S, Isshiki H, Sugita T, et al. A nuclear factor for IL-6 expression (NF-IL6) is a member of a C/EBP family. EMBO J. 1990;9:1897-1906.
69. Poli V, Mancini FP, Cortese R. IL-6DBP, a nuclear protein involved in interleukin-6 signal transduction, defines a new family of leucine zipper proteins related to C/EBP. Cell 1990;63:643-653.
70. Wegenka UM, Buschmann J, Lutticken C, Heinrich PC, Horn F. Acute-phase response factor, a nuclear factor binding to acute-phase response elements, is rapidly activated by interleukin-6 at the posttranslational level. Mol. Cell. Biol. 1993;13:276-288.
71. Nakajima K, Kusafuka T, Takeda T, Fujitani Y, Nakae K, Hirano T. Identification of a novel interleukin-6 response element containing an Ets-binding site and a CRE-like site in the junB promoter. Mol. Cell. Biol 1993;13:3027-3041.
72. Kojima H, Nakajima K, Hirano T. IL-6-inducible complexes on an IL-6 response element of the junB promoter contain Stat3 and 36 kDa CRE-like site binding protein(s). Oncogene 1996;12:547-554.
73. Zhang X, Blenis J, Li HC, Schindler C, Chen -KS. Requirement of serine phosphorylation for formation of STAT-promoter complexes. Science 1995;267:1990-1994.
74. Wen Z, Zhong Z, Darnell JE. Maximal activation of transcription by Stat1 and Stat3 requires both tyrosine and serine phosphorylation. Cell 1995; 82:241-250.
75. David M, Petricoin Er, Benjamin C, Pine R, Weber MJ, Larner AC. Requirement for MAP kinase (ERK2) activity in interferon alpha- and interferon beta-stimulated gene expression through STAT proteins [see comments]. Science 1995;269:1721-1723.
76. Pfeffer LM, Mullersman JE, Pfeffer SR, Murti A, Shi W, Yang CH. Stat3 as an adapter to couple phosphatidylinositol 3-kinase to the IFNAR1 chain of the type I interferon receptor. Science 1997;276:1418-1420.
77. Sekimoto T, Nakajima K, Tachibana T, Hirano T, Yoneda Y. Interferon-gamma-dependent nuclear import of Stat1 is mediated by the GTPase activity of Ran/TC4. J Biol Chem 1996;271:31017-31020.
78. Boulton TG, Stahl N, Yancopoulos GD. Ciliary neurotrophic factor/leukemia inhibitory factor/interleukin 6/oncostatin M family of cytokines induces tyrosine phosphorylation of a common set of proteins overlapping those induced by other cytokines and growth factors. J Biol Chem 1994;2696:11648-11655.
79. Fuhrer DK, Feng GS, Yang YC. Syp associates with gp130 and Janus kinase 2 in response to interleukin-11 in 3T3-L1 mouse preadipocytes. J. Biol. Chem 1995;270:24826-24830.
80. Fukada T, Hibi M, Yamanaka Y, et al. Two signals are necessary for cell proliferation induced by a cytokine receptor gp130: involvement of STAT3 in anti-apoptosis. mamuscript submitted. Immunity 1996;5:449-460.
81. Nakafuku M, Satoh T, Kaziro Y. Differentiation factors, including nerve growth factor, fibroblast growth factor, and interleukin-6, induce an accumulation of an active Ras.GTP complex in rat pheochromocytoma PC12 cells. J. Biol. Chem 1992;267:19448-19454.
82. Yin T, Yang YC. Mitogen-activated protein kinases and ribosomal S6 protein kinases are involved in signaling pathways shared by interleukin-11, interleukin-6, leukemia inhibitory factor, and oncostatin M in mouse 3T3-L1 cells. J. Biol. Chem. 1994;269:3731-3738.
83. Kumar G, Gupta S, Wang S, Nel AE. Involvement of Janus kinases, p52shc, Raf-1, and MEK-1 in the IL-6-induced mitogen-activated protein kinase cascade of a growth-responsive B cell line. J. Immunol 1994;153:4436-4447.
84. Berger LC, Hawley RG. Interferon-beta interrupts interleukin-6-dependent signaling events in myeloma cells. Blood 1997;89:261-271.
85. Daeipour M, Kumar G, Amaral MC, Nel AE. Recombinant IL-6 activates p42 and p44 mitogen-activated protein kinases in the IL-6 responsive B cell line, AF-10. J-Immunol 1993;150:4743-4753.
86. Ernst M, Oates A, Dunn AR. Gp130-mediated signal transduction in embryonic stem cells involves activation of Jak and Ras/mitogen-activated protein kinase pathways. J. Biol. Chem 1996;271:30136-30143.
87. Ernst M, Gearing DP, Dunn AR. Functional and biochemical association of Hck with the LIF/IL-6 receptor signal transducing subunit gp130 in embryonic stem cells. EMBO J. 1994;13:1574-1584.
88. Matsuda T, Takahashi Tezuka M, Fukada T, et al. Association and activation of Btk and Tec tyrosine kinases by gp130, a signal transducer of the interleukin-6 family of cytokines. Blood 1995;85:627-633.
89. Matsuda T, Fukada T, Takahashi Tezuka M, et al. Activation of Fes tyrosine kinase by gp130, an interleukin-6 family cytokine signal transducer, and their association. J. Biol. Chem. 1995;270:11037-11039.
90. Takahashi-Tezuka M, Hibi M, Fujitani Y, Fukada T, Yamaguchi T, Hirano T. Tec tyrosine kinase links the cytokine receptors to PI-3 kinase probably through JAK. Oncogene 1997;14:2273-2282.
91. Fukunaga R, Ishizaka Ikeda E, Pan CX, Seto Y, Nagata S. Functional domains of the granulocyte colony-stimulating factor receptor. EMBO J. 1991;10:2855-2865.
92. Murakami M, Narazaki M, Hibi M, et al. Critical cytoplasmic region of the interleukin 6 signal transducer gp130 is conserved in the cytokine receptor family. Proc Natl Acad Sci USA 1991;88:11349-11353.
93. Narazaki M, Witthuhn BA, Yoshida K, et al. Activation of JAK2 kinase mediated by the interleukin 6 signal transducer gp130. Proc Natl Acad Sci USA 1994;9181(6470):2285-2289.
94. Stahl N, Farruggella TJ, Boulton TG, Zhong Z, Darnell JE, Jr., Yancopoulos GD. Choice of STATs and other substrates specified by modular tyrosine-based motifs in cytokine receptors. Science 1995;267(5202):1349-1353.
95. Holgado-Madruga M, Emlet DR, Mosacatello DK, Godwin AK, Wang AJ. Grb2-associated docking protein in EGF- and insulin-receptor signalling. Nature 1996;319:560-564.
96. Weidner KM, Di-Cesare S, Sachs M, Brinkmann V, Behrens J, Birchmeier W. Interaction between Gab1 and c-Met receptor tyrosine kinase is responsible for epithelial morphogenesis. Nature 1996;384:173-176.
97. Keegan AD, Nelms K, White M, Wang L-M, Pierce JH, E. PW. An IL-4 receptor regin containing an insulin receptor motif is important for IL-4-mediated IRS-1 phosphorylation and cell growth. Cell 1994;76:811-820.
98. Johnston JA, Wang LM, Hanson EP, et al. Interleukin 2, 4, 7, and 15 stimulate tyrosine phosphorylation of insuline receptor substrates 1 and 2 in T cells. Potential role of JAK kinases. J. Biol. Chem. ;270:28527-28530.
99. Burfoot MS, Rogers NC, Watling D, et al. JAK-dependent activation of IRS-1 in response to IL-4, Oncostatin M and the interferons. J. Biol. Chem. 1997.
100. Yin T, Keller SR, Quelle FW, et al. Interleukin-9 induces tyrosine phosphorylation of insulin receptor substrate-1 via JAK tyrosine kinases. J Biol Chem 1995;270:20497-20502.
101. Argetsinger LS, Norstedt G, Billestrup N, White MF, Carter Su C. Growth hormone, interferon-gamma, and leukemia inhibitory factor utilize insulin receptor substrate-2 in intracellular signaling. J Biol Chem 1996;271:29415-29421.
102. Yamanaka Y, Nakajima K, Fukada T, Hibi M, Hirano T. Differentiation and growth arrest signals are generated through the cytoplasmic region of gp130 that is essential for Stat3 activation. EMBO J. 1996;15(7):1557-1565.
103. Baumann H, Symes A, J., Comeau MR, et al. Multiple regions within the cytoplasmic domains of the leukemia inhibitory factor receptor and gp130 cooperate in signal transduction in hepatic and neuronal cells. Mol. Cell. Biol. 1994;14:138-146.
104. Rincon M, Anguita J, Nakamura T, Fikrig E, Flavell RA. Interleukin (IL)-6 directs the differentiation of IL-4-producing CD4+ T cells. J. Exp. Med 1997;185:461-469.
105. Shabo Y, Lotem J, Rubinstein M, et al. The myeloid blood cell differentiation-inducing protein MGI-2A is interleukin 6. Blood 1988;72:2070-2073.
106. Miyaura C, Onozaki K, Akiyama Y, et al. Recombinant human interleukin 6 (B-cell stimulatory factor 2) is a potent inducer of differentiation of mouse myeloid leukemia cells (M1). FEBS Lett 1988;234:17-21.
107. Oritani K, Kaisho T, Nakajima K, Hirano T. Retinoic acid inhibits interleukin-6-induced macrophage differentiation and apoptosis in a murine hematopoietic cell line, Y6. Blood 1992;80:2298-2305.
108. Satoh T, Nakamura S, Taga T, et al. Induction of neuronal differentiation in PC12 cells by B-cell stimulatory factor 2/interleukin 6. Mol. Cell. Biol. 1988;8:3546-3549.
109. Nordan RP, Potter M. A macrophage-derived factor required by plasmacytomas for survival and proliferation in vitro. Science 1986;233:566-569.
110. Kawano M, Hirano T, Matsuda T, et al. Autocrine generation and requirement of BSF-2/IL-6 for human multiple myelomas. Nature 1988;332:83-85.
111. Van Snick J, Cayphas S, Szikora JP, et al. cDNA cloning of murine interleukin-HP1:homology with human interleukin 6. Eur J. Immunol 1988;18:193-197.
112. Hirano T. Interleukin 6 (IL-6) and its receptor: their role in plasma cell neoplasias. Int. J. Cell Cloning 1991;9:166-184.
113. Nakajima K, Yamanaka Y, Nakae K, et al. A central role for Stat3 in IL-6-induced regulation of growth and differentiation in M1 leukemia cells. EMBO J 1996;15:3651-3658.
114. Minami M, Inoue M, Wei S, et al. STAT3 activation is a critical step in gp1430-mediated terminal differentiation and growth arrest of a myeloid cell line. Proc. Natl. Acd. Sci. USA. 1996;93:3963-3966.
115. Shimozaki K, Nakajima K, Hirano T, Nagata S. Involvement of STAT3 in the granulocyte colony stimulating factor-induced differentiation of myeloid cells. J. Biol. Chem. 272:25184-25189, 1997
116. Chin YE, Kitagawa M, Su WC, You ZH, Iwamoto Y, Fu XY. Cell growth arrest and induction of cyclin-dependent kinase inhibitor p21 WAF1/CIP1 mediated by STAT1. Science 1996;272:719-722.
117. Morse L, Chen D, Franklin D, Yue Xiong, Chen-Kiang S. Induction of Cell Cycle Arrest and B Cell Terminal Differentiation
by CDK Inhibitor p18INK4c and IL-6. Immunity 1997;6:47-56.
118. Billadeau D, Jelinek DF, Shah N, LeBien TW, Van Ness B. Introduction of an activated N-ras oncogene alters the growth characteristics of the interleukin 6-dependent myeloma cell line ANBL6. Cancer Res 1995;55:3640-3646.
119. Schwarze MM, Hawley RG. Prevention of myeloma cell apoptosis by ectopic bcl-2 expression or interleukin 6-mediated up-regulation of bcl-xL. Cancer Res 1995;55:2262-2265.
120. Wu YY, Bradshaw RA. Induction of neurite outgrowth by interleukin-6 is accompanied by activation of Stat3 signaling pathway in a variant PC12 cell (E2) line. J. Biol. Chem 1996;271:13023-13032.
121. Ihara S, Iwamatsu A, Fujiyoshi T, Komi A, Yamori T, Fukui Y. Identification of interleukin-6 as a factor that induces neurite outgrowth by PC12 cells primed with NGF. J. Biochem 1996;120:865-868.
122. Sterneck E, Kaplan DR, Johnson PF. Interleukin-6 induces expression of peripherin and cooperates with Trk receptor signaling to promote neuronal differentiation in PC12 cells. J Neurochem 1996;67:1365-1374.
123. Ihara S, Nakajima K, Fukada T, et al. Dual control of neurite outgrowth by STAT3 and MAP kinase in PC12 cells stimulated with interleukin-6. EMBO J. 16:5345-5352, 1997
124. Boeuf H, Hauss C, De Graeve F, Baran N, Kedinger C. LIF-dependent transcriptional activation in embryonic stem cells. J. Cell. Biology 138:1207-1217, 1997
125. Ihle JN. STATs: Signal transducers and Activators of Transcription. Cell 1996;84:331-334.
126. O'Shea JJ. Jaks, STATs, cytokine signal transduction, and immunoregulation: are we there yet ? Immunity 1997;7:1-11.
127. Liu ZG, Hsu H, Goeddel DV, Karin M. Dissection of TNF receptor 1 effector functions: JNK activation is not linked to apoptosis while NF-kappaB activation prevents cell death. Cell 1996;87:565-576.
128. Qui MS, Green SH. PC12 cell neuronal differentiation is associated with prolonged p21ras activity and consequent prolonged ERK activity. Neuron 1992;9:705-717.
129. Traverse S, Gomez N, Paterson H, Marshall C, Cohen P. Sustained activation of the mitogen-activated protein (MAP) kinase cascade may be required for differentiation of PC12 cells. Comparison of the effects of nerve growth factor and epidermal growth factor. Biochem J. 1992;288:351-355.
130. Haspel RL, Salditt Georgieff M, Darnell JE, Jr. The rapid inactivation of nuclear tyrosine phosphorylated Stat1 depends upon a protein tyrosine phosphatase. EMBO J. 1996;15:6262-6268.
131. Caldenhoven E, van Dijk TB, Solari R, et al. STAT3beta, a splice variant of transcription factor STAT3, is a dominant negative regulator of transcription. J. Biol. Chem 1996;271:13221-13227.
132. Wang D, Stravopodis D, Teglund S, Kitazawa J, Ihle JN. Naturally occurring dominant negative variants of Stat5. Mol. Cell. Biol 1996;16:6141-6148.
133. Klingmuller U, Lorenz U, Cantley LC, Neel BG, Lodish HF. Specific recruitment of SH-PTP1 to the erythropoietin receptor causes inactivation of JAK2 and termination of proliferative signals. Cell 1995;80:729-738.
134. Ali S, Chen Z, Lebrun JJ, et al. PTP1D is a positive regulator of the prolactin signal leading to beta-casein promoter activation. EMBO J. 1996;15:135-142.
135. Kharitonenkov A, Chen Z, Sures I, Wang H, Schilling J, Ullrich A. A family of proteins that inhibit signalling through tyrosine kinase receptors. Nature 1997;386:181-186.
136. Fujioka Y, Matozaki T, Noguchi T, et al. A novel membrane glycoprotein, SHPS-1, that binds the SH2-domain-containing protein tyrosine phosphatase SHP-2 in response to mitogens and cell adhesion. Mol. Cell. Biol 1996;16:6887-6899.
137. Starr R, Willson TA, Viney EM, et al. A family of cytokine-inducible inhibitors of signalling. Nature 1997;387:917-921.
138. Endo TA, Masuhara M, Yokouchi M, et al. A new protein containing an SH2 domain that inhibits JAK kinases. Nature 1997;387:921-924.
139. Naka T, Narazaki M, Hirata M, et al. Structure and function of a new STAT-induced STAT inhibitor. Nature 1997;387:924-929.
ü@
|
|
|