|
|
|
Ten years have passed since the molecular cloning of interleukin 6 (IL-6) in 1986. IL-6 is a typical cytokine, exhibiting functional pleiotropy and redundancy. IL-6 is involved in the immune response, inflammation, and hematopoiesis. The IL-6 receptor consists of an IL-6 binding a chain and a signal transducer, gp130, which is shared among the receptors for the IL-6 related cytokine subfamily. The sharing of a receptor subunit is a general feature of cytokine receptors and provides the molecular basis for the functional redundancy of cytokines. JAK tyrosine kinase is a key molecule that can initiate multiple signal-transduction pathways by inducing the tyrosine-phosphorylation of the cytokine receptor, gp130 in the case of IL-6, on which several signaling molecules are recruited, including STAT, a signal transducer and activator of transcription, and SHP-2, which links to the Ras-MAP kinase pathway. JAK can also directly activate signaling molecules such as STAT and Tec. These multiple signal-transduction pathways intimately regulate the expression of several genes including c-myc, c-myb, junB, IRF1, egr-1, and bcl-2, leading to the induction of cell growth, differentiation, and survival. The deregulated expression of IL-6 and its receptor is involved in a variety of diseases. (to The table of contents)
Ten years have passed since interleukin 6 (IL-6) was cloned in 1986 (1-4). During the last decade, many findings were made concerning the structure and function of IL-6 and its receptor, and the role of IL-6 in a variety of diseases. These findings together with a large volume of studies on many cytokines have greatly contributed to the establishment of a variety of concepts about cytokines in general: the establishment of pleiotropy and redundancy as properties of cytokine function, the cytokine receptor super family, the sharing of a signal-transducing receptor subunit among several cytokine receptors, the agonistic activity of certain soluble cytokine receptors, and the elucidation of the molecular mechanisms of inflammatory diseases.
In early 1980, Muraguchi and Kishimoto and their colleagues (5) and Teranishi and Hirano and their colleagues (6) independently showed that the culture supernatant fractions of stimulated human peripheral blood mononuclear cells and tonsillar mononuclear cells, respectively, induce immunoglobulin (Ig) production in Epstein-Barr virus (EBV)-transformed B lymphoblastoid cell lines. Kishimoto's group called this active factor, "TRF" or "BCDF" (5,7) and Hirano's group termed it, "TRF-like factor" or "BCDFII"(6,8,9). Teranishi et al. (6) partially purified the TRF-like factor and showed that it is recovered in the fractions corresponding to molecular weights of 22 kDa and 36 kDa, by gel filtration, and that its isoelectric point is 5 to 6. This active factor was further purified to homogeneity, renamed "BSF-2 "(10), and molecularly cloned (1). The molecular cloning of interferon (IFN) b2 (2,3), 26kDa protein (4), and BSF-2 (1), independently achieved in 1986, revealed that all these molecules are identical (11,12). Furthermore, the plasmacytoma/hybridoma/myeloma growth factor (13-17) and the hepatocyte stimulating factor (18,19), which regulates the biosynthesis of a variety of acute-phase proteins, were also found to be identical to this factor. Therefore, the name, "interleukin 6" has been applied to this molecule (20). IL-6 is now known to be a multifunctional cytokine that plays roles in the immune response, inflammation, hematopoiesis, and in the endocrine and nervous systems (12,20-24).
Many cytokines possess a similar helical structure (25,26). The IL-6 receptor and many other cytokine receptors are also structurally similar and constitute the cytokine receptor super family (25,27,28). In addition, cytokine receptor subunits are shared among several cytokine receptors (24,29-33). This sharing of subunits is one of the mechanisms by which the functional redundancy of cytokine activities occurs. A typical example is the gp130 subunit, which is shared among the receptors for IL-6, leukemia inhibitory factor (LIF), ciliary neurotrophic factor (CNTF), oncostatin M (OSM), IL-11, and cardiotrophin-1 (CT-1) (24,32-40). Studies on the signal transduction of IFNs have shown that novel tyrosine kinases, JAKs (Janus kinases), and transcriptional factors, STATs (signal transducer and activator of transcription), play a major role in signal transduction through the receptors for a variety of cytokines and hormones (31-33,41-44). The JAK-STAT pathway seems to play essential role in cytokine function and in the expression of the functional specificity of each cytokine.
In this review, I describe the recent progress in studies of the biology of IL-6 and of signal transduction through the IL-6 receptor, with emphasis on the molecular mechanisms involved in the regulation of growth and differentiation through the IL-6 receptor.(to The table of contents)
IL-6 is a typical example of a pleiotropic cytokine that acts on various cells: IL-6 induces the differentiation of B cells to antibody producing plasma cells, T-cell growth and differentiation, the differentiation of myeloid leukemic cell lines into macrophages, megakaryocyte maturation, the neural differentiation of PC12 cells, the development of osteoclasts, and acute-phase protein synthesis in hepatocytes. IL-6 acts as a growth factor for myeloma/plasmacytoma, keratinocytes, mesangial cells, renal cell carcinoma, and Kaposi's sarcoma, and promotes the growth of hematopoietic stem cells. In addition, IL-6 inhibits the growth of myeloid leukemic cell lines and certain carcinoma cell lines (see reviews 12, 20-24). In accordance with the multiple functions of IL-6, IL-6 transgenic mice or mice bearing a retrovirus vector expressing IL-6 show massive plasmacytosis, hypergammaglobulinemia, an increase in acute-phase proteins, mesangial proliferative glomerulonephritis, an increase in megakaryocytes (45,46), and eventually develop plasmacytoma (47). Based on the observations using IL-6 transgenic mice, together with the functional activity of IL-6 as a growth factor of myeloma and plasmacytoma, it was hypothesized that the deregulated expression of the IL-6 gene is involved in the generation of pristane-induced murine plasmacytoma and plays an important role in the development of myeloma in humans (48). In fact, it was shown using IL-6-deficient mice that IL-6 is essential for the development of plasmacytoma in vivo (49). IL-6 deficient mice showed a reduced IgG response, but no reduction in the IgM response to both a soluble protein antigen and vesicular stomatitis virus (VSV) antigen (50). A striking effect was observed in the mucosal IgA antibody response. In accordance with the original findings of Kiyono and McGhee and their colleagues (51,52) that IL-6 is involved in the IgA response, in IL-6 deficient mice, the number of IgA-producing cells was greatly reduced (53). This reduced IgA response was completely restored after intranasal infection with recombinant vaccinia viruses engineered to express IL-6. The generation of cytotoxic T cells against vaccinia virus was 3- to 10-fold reduced in IL-6 deficient mice, while CTL function against lymphocytic choriomeningitis virus (LCMV) was not reduced (50). Furthermore, the inflammatory acute-phase response after tissue damage or infection is severely compromised (50). An inability to clear Listeria monocytogenes was observed in IL-6 deficient mice (50,54). This inability is most likely due to the inability of neutrophils to function in IL-6 deficient mice, suggesting that IL-6 plays a critical role in listeriosis by stimulating neutrophils (54). Since an anti-IL-6 antibody can inhibit the increase in osteoclast precursors occurring in estrogen-depleted mice (55), and since estrogen can inhibit the IL-1- and TNFa-induced production of IL-6 (56), the overproduction of IL-6 has been suggested to be involved in the generation of postmenopausal osteoporosis. In support of this notion, ovariectomy does not induce any change in either bone mass or bone remodeling rates in IL-6 deficient mice, although estrogen deficiency induced by ovariectomy causes a significant loss of bone mass together with an increase in bone turnover rates in wild type mice (57). Intraperitoneal injections of either LPS or IL-1 beta failed to evoke a fever response in IL-6 deficient mice and the fever response was recovered by the intracerebroventricular injection of recombinant human IL-6, but not of IL-1, showing that IL-6 is a necessary component of the fever response to both IL-1 and LPS (58). These facts show that IL-6 is critical in only a limited range of biological reactions, such as the acute-phase response, the mucosal IgA response, the fever response, and estrogen deficiency-induced bone loss, although IL-6 has many biological activities. The biological activities of IL-6 may be compensated for by other cytokines showing functional redundancy with IL-6.(to The table of contents)
Functional redundancy among the IL-6 family cytokines
IL-6, LIF, CNTF, OSM, IL-11, and CT-1 constitute the IL-6 related cytokine subfamily because of their functional redundancy, structural similarity, and sharing of a receptor subunit.
LIF has many biological activities (59,60), including the induction of monocytic differentiation in a murine leukemic cell line, M1, the suppression of the differentiation of pluripotent embryonic stem (ES) cells (61), and the inhibition of adipogenesis (62). Furthermore, LIF has been shown to be identical to the cholinergic neuronal differentiation factor (63) that controls neuronal phenotypic decisions. CNTF was characterized as a survival factor for chick ciliary neurons in vivo (64,65). CNTF promotes the survival of a variety of other neuronal cell types (66,67) and affects the differentiation of E7 chick sympathetic neurons (68). CNTF also induces the cholinergic differentiation of sympathetic neurons from newborn rats (69). Moreover, CNTF promotes the differentiation of bipotential O-2A progenitor cells to type-2-astrocytes in vitro (70). Thus, both CNTF and LIF promote the transition from noradrenergic to cholinergic function in cultured sympathetic neurons. In addition, both CNTF and LIF affect the survival and differentiation of motor and sensory neurons (71,72). OSM was identified by its ability to inhibit the growth of human melanoma cell line A375 (73,74). OSM is also a potent growth factor for AIDS-related Kaposi's sarcoma (AIDS-KS) (75,76) and enhances the secretion of IL-6 by AIDS-KS cells. IL-11 was identified as a growth factor for a mouse plasmacytoma cell line (77) and found to augment antigen-specific antibody responses (78). CT-1 is a cytokine that induces cardiac myocyte hypertrophy (40). CT-1 inhibits the growth of M1 leukemia cells, induces a phenotypic switch in sympathetic neurons, and promotes the survival of rat dopaminergic and chick ciliary neurons (79).
These facts illustrate the functional redundancy that occurs among these cytokines. In particular, IL-6, LIF, OSM, and CT-1 induce macrophage differentiation in a myeloid leukemic cell line, M1(23,24,80,81). IL-6, IL-11, LIF, and OSM all induce the growth of myeloma cells (17,82). IL-6, LIF, and IL-11 enhance IL-3-dependent colony formation of primitive blast colony-forming cells (23,24,83,84). IL-6, LIF, IL-11, and OSM stimulate the biosynthesis of acute-phase proteins in hepatocytes (22,85-88). IL-6, LIF, OSM, IL-11, CNTF, and CT-1 all act on the nervous system (89). The functional redundancy observed among the IL-6-related cytokine subfamily is largely explained by the sharing of the receptor subunit, gp130.(to The table of contents)
IL-6 receptor system and the sharing of gp130 among the receptors for the IL-6 related cytokine subfamily
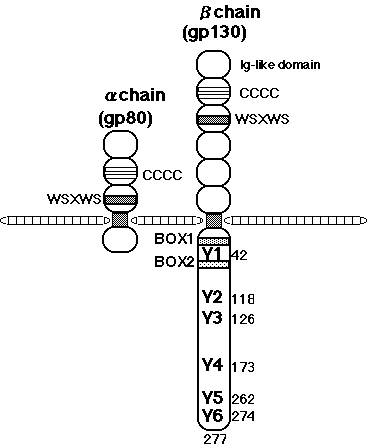
The IL-6 receptor (R) complex consists of an 80 kDa IL-6 binding molecule termed IL-6Ra, and a signal transducer, gp130 (Figure 1) (34,35,90) Many cytokine receptors are similar in structure to IL-6Ra and constitute the type I cytokine receptor super family (25,28). The most striking features of these receptors are the conservation of four cysteine residues and a tryptophan-serine-X-tryptophan-serine (W-S-X-W-S) motif (WS motif) located just outside the transmembrane domain (Figure 1).
The cytoplasmic domain of IL-6Ra is not necessary for signal transduction (34,91). Furthermore, the soluble form of IL-6Ra acts in an agonistic, rather than an antagonistic manner. These observations led to the discovery of gp130, a signal transducing subunit of the IL-6R system. The binding of IL-6 to its receptor triggers the association of IL-6Ra with another membrane-associated 130kD glycoprotein, gp130 (34). The cDNA encoding gp130 was cloned and gp130 was found to belong to the cytokine receptor super family (35). By itself, gp130 does not bind IL-6, but IL-6Ra together with gp130 form a high affinity IL-6 binding site. The complex composed of IL-6 and soluble IL-6Ra associates with gp130 and generates growth signals in the cells that express gp130. The complete structure of the IL-6R system was recently found to be a hexamer composed of two each of IL-6, IL-6R, and gp130 ( ). Most importantly, it was found that gp130 is shared among the receptors for CNTF, LIF, OSM, IL-11, and CT-1, and that it is critically involved in generating signal transduction through these receptors (24,32,33).
The sharing of a receptor subunit among different cytokine receptors has also been demonstrated. Granulocyte-macrophage colony stimulating factor (GM-CSF), IL-3, and IL-5 receptors share a common b subunit (29). The b chain of the IL-2 receptor (IL-2Rbc, c for common) is shared by the IL-15 receptor, and the g chain of the IL-2 receptor (gc) is shared by the IL-4, IL-7, IL-9, and IL-15 receptors (30,31). Thus, the molecular mechanisms of redundancy in cytokine activity could be explained at least in part by the sharing of receptor subunits among several cytokine receptors (Figure 2) .(to The table of contents)
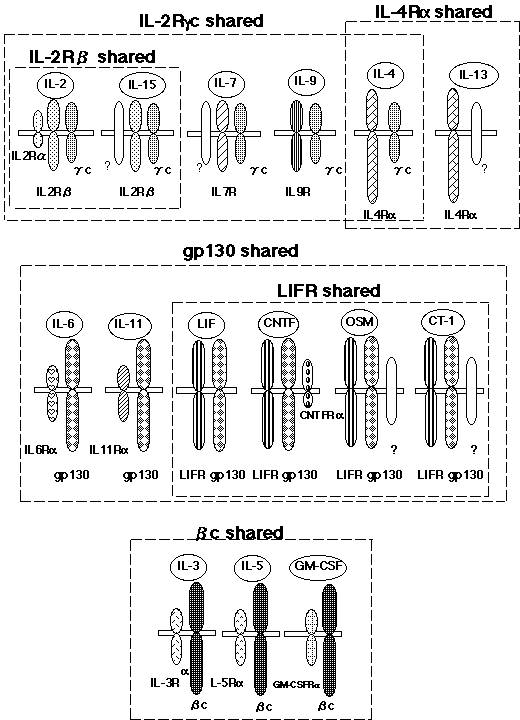
A novel mechanism generating cytokine diversity.(see News and Topics)
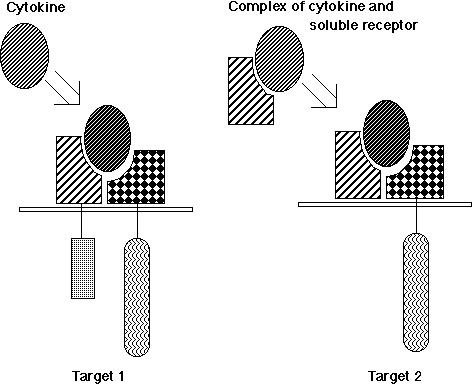
Investigations of the IL-6R system have provided evidence that a complex of IL-6 and a soluble form of IL-6Ra could act on the cells that express gp130, but not the a-chain. This kind of model can apply to other cytokine receptor systems. IL-12 consists of a disulfide heterodimer of 40kD (p40) and 35kD (p35) subunits (93). The peptide sequences of p35 and p40 resemble IL-6 and the soluble form of its receptor, respectively (94), suggesting that IL-12 acts on target cells in a manner similar to the complex of IL-6 and soluble IL-6R. This model is also supported by the fact that molecular cloning of IL-12R showed that it is a member of the cytokine receptor super family, most closely related to gp130 (95). Another example is a CNTFRa that is anchored to the cell membrane by a glycosyl-phosphatidyl inositol (GPI) linkage. The complex of soluble CNTFRa and CNTF acts on the cells that express LIFRb and gp130 (96). Potential physiological roles for the soluble CNTFRa are suggested by the presence of the soluble form of the a-chain in cerebrospinal fluid and its release from skeletal muscle in response to peripheral nerve injury.
Based on these facts, I originally proposed a novel mechanism by which the cytokine system generates functional diversity (Figure 3) (24). A complex consisting of a soluble cytokine receptor and its corresponding cytokine acquires a different target specificity from the original cytokine and, therefore, it should express functions distinct from the original cytokine. Actually, double transgenic mice expressing human IL-6 and IL-6Ra showed myocardial hypertrophy (97), indicating that the complex of IL-6 and the soluble form of IL-6Ra acts on heart muscle cells that express gp130, on which IL-6 alone cannot act, leading to the induction of cardiac hypertrophy similar to the effect of CT-1. This model could also be applied to the glial-cell-line-derived neurotrophic factor (GDNF) receptor system, which consists of a GDNF-specific binding molecule, GDNFRa, which is a GPI-anchored membrane molecule, and a signal transducing GDNFR, Ret, which is a receptor tyrosine kinase (98,99). I anticipate that this novel mechanism will be applied to a wide range of other receptor systems. This mechanism may contribute to generating the functional diversity of cytokines and may also play pathological roles in various diseases, since an increase in the serum-soluble form of various cytokine receptors has been reported to occur in a variety of diseases.(to The table of contents)
Signal transduction through gp130: JAK as a key molecule initiating signaling and STAT as a major signal transducer through the cytokine receptor
Since the cytoplasmic domain of most cytokine receptors, including gp130, does not have an intrinsic catalytic domain, one of the hottest issues until 1993 was the identification of catalytic molecules that associate with cytokine receptors and can be activated by cytokines. This issue was resolved by the findings that Tyk2, a JAK family tyrosine kinase, is critical for IFNa signaling (100), and that JAK2 is associated with the erythropoietin receptor and the growth-hormone receptor, and is activated by these ligands(101,102). Since then, several JAK family tyrosine kinases (JAK1, JAK2, JAK3, Tyk2) have been shown to be involved in the signal transduction of cytokines and hormones, and to function as a central non-receptor tyrosine kinase that can initiate cytokine signaling (41). JAK (Janus kinase, or just another kinase) has a kinase domain (JH1, JH; JAK homology), a kinase-like domain (JH2) and several domains that are conserved among JAK family tyrosine kinases (JH3-JH7), but does not have any SH2 or SH3 domains. Furthermore, the signal transducer and activator of transcription (STAT) , which was originally identified as an interferon-activated transcription factor by Darnell and his colleagues, has been found to be involved in a variety of cytokine signal-transduction pathways (42,43).
JAK1, JAK2, and Tyk2 have been shown to associate constitutively with gp130 and to be tyrosine-phosphorylated in response to IL-6, CNTF, LIF, or OSM (103-105). Furthermore, IL-6 has been shown to activate STAT3, STAT1, and very recently, STAT5 (106-111). Two types of IL-6 responsive elements (REs) have been identified in the genes encoding acute-phase proteins. Type I IL-6 RE, which is a binding site for NF-IL6 (112), is present in the CRP, hemopexin A, and haptoglobin genes. The binding activity of NF-IL6 is most likely induced by IL-6 through the increased expression of the NF-IL6 gene, rather than through its posttranslational modification (113,114). The type II IL-6 RE is found in the fibrinogen, a2 macroglobulin, a1 acid glycoprotein, and haptoglobin genes. It has been shown that IL-6 triggers the rapid activation of a nuclear factor, termed acute-phase response factor (APRF), which binds to type II IL-6 RE (115). The purification and molecular cloning of APRF revealed that it is identical with STAT3 (106,107). In parallel with these studies, Nakajima et al. (116) identified the IL-6 responsive element of the junB gene (JRE-IL6). This element consists of a putative Ets binding site (JEBS) and CRE-like site. Both sites are essential for the JRE-IL6 to be transcriptionally activated. Furthermore, the IL-6-inducible JEBS-binding protein mainly contains STAT3, although the JEBS is a low-affinity binding site for STAT3 as compared with APRE (108,109). IL-6 induces the formation of a complex consisting of STAT3 and p36-CRE-like site binding molecules on the JRE-IL6, as well as on the IL-6/IFNg responsive element in the IRF-1 promoter (117). Such binding complex formation seems to be important for STAT to act on a low-affinity binding site, such as the JEBS, and therefore may contribute to generating the diversity of target genes of STAT proteins. In addition to the tyrosine phosphorylation of STAT3 by JAK tyrosine kinase, we showed that the H7-sensitive pathway, most likely a serine/threonine kinase, is required for both the JRE-IL6 and STAT3 to be transcriptionally active (109, 116), although H7 did not inhibit the IL-6-induced appearance of the DNA binding complex on the APRE or JEBS (109). In certain cell lines, it has been shown that STAT3 requires phosphorylation on serine to form a STAT3-STAT3 homodimer, which binds to DNA (118), further suggesting the involvement of a serine/threonine kinase in the STAT signal pathway. In fact, maximal activation of transcription by STAT1 and STAT3 requires both tyrosine and serine phosphorylation (119).(to The table of contents)
Multiple signal-transduction pathways through gp130 regulating cell growth, differentiation, and survival
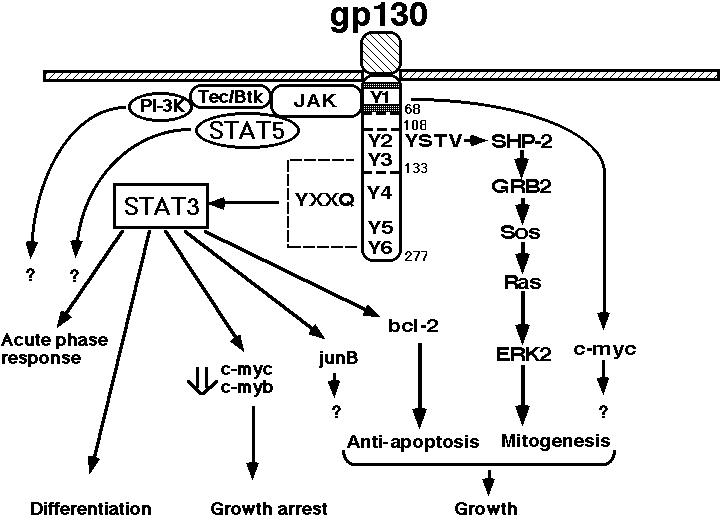
Human gp130 has 277 amino acid residues in its cytoplasmic domain, which contains two motifs conserved among the cytokine receptor family, termed box 1 and box 2 (Fig. 1 and Fig. 4) (35,120,121). The membrane-proximal region containing box1 and box2 is sufficient for JAK to be activated through gp130 (122). Gp130 has six tyrosine residues in its cytoplasmic domain, and the tyrosine phosphorylation of SHP-2 (also called PTP1-D, SHPTP-2, PTP2C, and Syp), a phosphotyrosine phosphatase, and that of STAT3 are dependent on the second tyrosine from the membrane (Y2), and any one of the four tyrosines (Y3, Y4, Y5, Y6) in the carboxy terminus that have a glutamine residue at the third position behind tyrosine (Y-X-X-Q), respectively (Fig. 4) (123,124). In accordance with the fact that STAT3 is involved in the activation of type II acute-phase genes, the membrane-proximal region of gp130, containing 133 amino acids and Y3, is necessary for the activation of IL-6 responsive acute-phase genes (125).
|
|
 |
|
|
|
IL-6 induces growth arrest and macrophage differentiation in the murine myeloid leukemic cell lines, M1 and Y6 (126,127,128). Yamanaka et al. (124) showed that the membrane-proximal region of gp130, consisting of 133 amino acids, is sufficient to generate the signals for growth arrest, macrophage differentiation, down-regulation of c-myc and c-myb, induction of junB and IRF1, and the activation of STAT3. The region between 108 and 133 amino acids contains two tyrosine residues (Fig. 1 and Fig. 4): one (Y3) at amino acid position 126 with the YXXQ motif, and the other one (Y2) without the motif at amino acid position 118. Y2 has been shown to be essential for gp130-mediated egr-1 gene induction (124). Yamanaka et al. (124) showed that Y3, which has the motif, was critical in generating the signals not only for STAT3 activation but also for growth arrest and differentiation, accompanied by the down-regulation of c-myc and c-myb and the immediate early induction of junB and IRF1. These results suggest that STAT3 plays an essential role in the signals for growth arrest and differentiation. This possibility was proven by Nakajima et al.(129), who introduced two types of mutant STAT3s acting in a dominant-negative manner into M1 leukemic cells. They showed that dominant-negative forms of STAT3 inhibited both IL-6-induced growth arrest and macrophage differentiation in the M1 transformants. Blocking the STAT3 activation resulted in the inhibition of IL-6-induced repression of c-myb and c-myc, but not egr-1 induction. Furthermore, IL-6 enhanced the growth of M1 cells when STAT3 was suppressed. Thus, IL-6 generates both growth-enhancing signals and growth arrest- and differentiation-inducing signals at the same time, but the former is only apparent when STAT3 activation is suppressed. The essential role of STAT3 in the IL-6-induced macrophage differentiation of M1 cells was also shown by Minami et al.(130).
For the growth signal, it was shown that a 65-amino acid region proximal to the transmembrane domain is sufficient for the growth response, by using gp130-transfectants of an IL-3-dependent Pro-B-cell line BAF/B03 (32,121). However, Fukada et al.(131) showed that the membrane-proximal region of 68 amino acids is not sufficient to induce 3H-Tdr uptake when cells are starved of IL-3, although this region is sufficient to induce c-myc gene activation. The membrane-proximal region containing 133 amino acid residues is required and sufficient for cell growth. Furthermore, they showed that at least two distinct signals are required for gp130-induced cell growth: one is a mitogenic signal dependent on the second tyrosine residue, Y2, and possibly mediated by SHP-2, and the other is an anti-apoptotic signal dependent on the third tyrosine residue, Y3, and mediated by STAT3: Mutations of Y2 abolish the tyrosine phosphorylation of SHP-2, the association between SHP-2 and GRB2, and the activation of MAP kinase. Mutations of Y2 result in the loss of gp130-mediated cell growth but still induces anti-apoptotic signals resulting in cell survival for at least four days. On the other hand, mutations of Y3 disrupt STAT3 activation and result in apoptosis and the loss of bcl-2 induction. Moreover, they showed that dominant-negative STAT3 inhibits both gp130-mediated anti-apoptotic signals and bcl-2 induction, indicating that STAT3 plays a central role in gp130-mediated anti-apoptotic signals.
Thus, STAT3 plays pivotal roles in gp130-mediated signal transduction regulating cell growth, differentiation, and survival. Furthermore, STATs play essential roles in a variety of biological functions; STAT1 is critical for interferons (IFNs) to exert their functions, and is also involved in innate immunity (132,133). STAT6 is essential for IgE class switching and the Th2 response in the immune system (134-136). STAT4 is critically involved in the IL-12 response and Th1 activation (137,138). D-STAT regulates the expression of pair-rule genes and is critically involved in Drosophila early development (139,140). Thus, the molecular mechanism by which STATs are selectively activated should give the molecular basis for the expression of the specific biological activity of each cytokine. Activated JAK phosphorylates specific tyrosine residues on gp130, thereby creating the docking sites that are specifically recognized by the SH2 domain of STAT3; STAT3 activation requires the phosphorylated tyrosine residues fitting a YXXQ consensus motif located on the distal domain of gp130 (123,124). Similar mechanisms have also been shown for the activation process of other STATs, for example, STAT1 by the IFNg receptor (141), STAT6 by the IL-4 receptor a chain (142), and STAT5 by the IL-2 receptor b chain and the erythropoietin receptor (143-146). These studies suggest that the activation of STATs strictly depends on the specific tyrosine residues on their corresponding receptors, although recent studies have indicated that in some cases STATs might be activated independently of the tyrosine residues on the receptors (110,147,148). Fujitani et al. (111) showed that STAT5 can be activated independently of tyrosine phosphorylation of gp130, and that its activation requires direct interaction with the JH2 domain of JAKs. These observations indicate that the activation of STATs involves various mechanisms, including the interaction between the JH2 domain of JAKs and STATs, or the SH2 domain of STATs and cytokine receptors (Figure 5). On the other hand, different species of JAK family tyrosine kinases are activated by a variety of cytokines. For example, Epo, growth hormone, and IL-3 activate only JAK2. IL-2 activates JAK1 and JAK3, which associate with IL-2Rb and IL-2Rg, respectively (149). The signaling pathway of IFNa and b requires both Tyk2 and JAK1, whereas JAK1 and JAK2 are activated and essential for the IFNg receptor-mediated signal pathway (150). However, in the absence of JAK1, transcriptional factor STATs (STAT1 and STAT3) are not activated efficiently upon stimulation by IL-6, although JAK2 and Tyk2 are activated, suggesting that there is a hierarchy among gp130-associated JAKs (151).
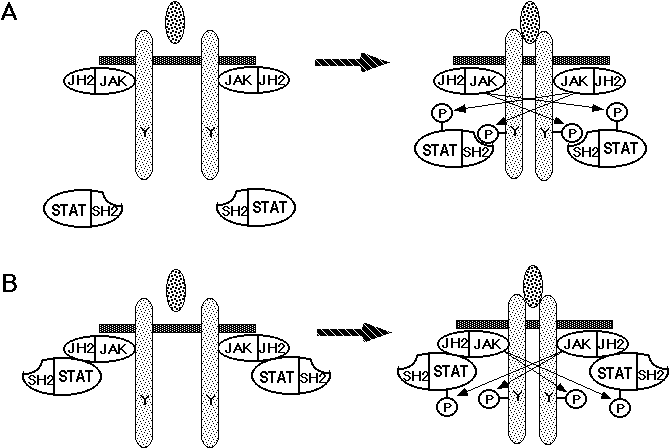
Taken together, all evidence indicates that the combination of the binding specificity of the SH2 domain of STAT for the tyrosine-phosphorylated cytokine receptor, that of the JH2 domain of JAK for STAT, the set of JAKs that associate with a given receptor, and the spectrum of these molecules expressed in a given cell, eventually determines the specific JAK-STAT signal-transduction pathway that is activated through a given cytokine receptor in a particular cell type, leading to the generation of specific biological activity through a cytokine receptor .
In addition to the JAK-STAT signal-transduction pathway, the Ras-MAP kinase pathway is activated through SHP-2 (131) or Shc(152). Furthermore, non-receptor tyrosine kinases, such as Btk, Tec, Fes, and Hck (153-155) are activated through the IL-6 receptor, as well as through a variety of other cytokine receptors (31). Takahashi-Tezuka et al.(156) showed that Tec associates with JAK1 and JAK2 and Btk associates with JAK1. Furthermore, both Tec and Btk associate with the p85 subunit of PI-3 kinase, and Tec binds to Vav, although the biological significance of these signal-transduction pathways remains to be elucidated. As summarized in Figure 4, several distinct signal-transduction pathways are generated through different regions of the cytoplasmic domain of gp130. The set of signaling pathways that are activated in a given cell may differ, depending on the expression pattern of these signaling molecules. Furthermore, these signaling pathways may interact with each other and contribute to a variety of biological activities.(to The table of contents)
IL-6 and B-cell abnormalities: Expression of IL-6 and its receptor in autoimmune disease.
We have suggested that IL-6 is involved in B-cell abnormalities in patients with cardiac myxoma who show hypergammaglobulinemia (157). We previously showed that the pleural effusion cells of patients with pulmonary tuberculosis produce a large amount of factors capable of inducing immunoglobulin production (158), and one of these active factors was partially purified and designated as TRF-like factor/IL-6 (6). It is noteworthy that patients with pulmonary tuberculosis often have a wide range of autoantibodies (159) and that in certain cases, a diffuse hypergammaglobulinemia has been observed (160). These facts suggest that IL-6 plays some roles in B-cell abnormalities associated with the inflammatory process. Consistent with this notion, it has been implicated that inflammation plays a role in polyclonal B-cell activation and monoclonal B-cell neoplasia. Patients with pre-existing chronic inflammations show polyclonal hypergammaglobulinemia and frequently develop plasma cell neoplasias or lymphoma (159,161-163). Mineral oil or pristane, which induces chronic inflammation, elicits not only an erosive arthritis resembling rheumatoid arthritis (RA) but also plasmacytoma in certain strains of mice (164,165). Furthermore, pristane induces the production of autoantibodies characteristic of systemic lupus erythematosus (SLE) (166). It is noteworthy that pristane is a strong inducer of IL-6 (16). Abnormal IL-6 production is observed in patients with RA (167-169). It has been demonstrated that there is a significant correlation between the concentrations of synovial fluid IL-6 and IgG in RA (170) . Significant correlation is also shown between serum IL-6 activity and serum levels of a variety of acute-phase proteins (168). T cells, B cells, synoviocytes and chondrocytes are identified as sources of IL-6 (167,171,172). IL-6 production is also observed in type II collagen-induced arthritis in mice (173) and MRL/lpr mice (174), which develop autoimmune disease with proliferative glomerulonephritis and arthritis.
Activated B cells, but not resting B cells, express the IL-6 receptor (175). Abnormal expression of the IL-6 receptor is shown in several autoimmune diseases. B cells in patients with SLE produce IL-6 and respond to IL-6, leading to the production of Ig, suggesting that SLE B cells constitutively express the IL-6 receptor (176,177). Constitutive expression of the IL-6 receptor in SLE patients has been proven by utilizing specific antibodies, and antibodies against the IL-6 receptor inhibit the spontaneous production of anti-DNA autoantibodies (178). Constitutive expression of the IL-6 receptor has also been demonstrated in splenic B cells from aged MRL/lpr mice (179). IL-6, but neither IL-2, IL-4, nor IL-5 enhanced the IgG anti-DNA antibody production by splenic B cells from NZB x NZW (NZB/W)F1 mice over 6 months of age. The ability of B cells to produce IgG anti-DNA antibodies seems to be related to the expression levels of IL-6 receptor at the age when the mice begin to develop the disease (180). Administration of IL-6 to (NZB x NZW)F1 mice results in an accelerated and severe form of membranoproliferative glomerulonephritis (181). IL-6 is produced by islet b-cells and the thyroid (182,183). By enhancing the response of auto-reactive T cells, IL-6 may be involved in type I diabetes, in which T cells are critical to the initiation and development of autoimmune b-cell destruction (184). Another interesting piece of evidence is that a striking increase in the level of agalactosyl IgG has been observed in a variety of autoimmune and/or IL-6 related diseases, such as pulmonary tuberculosis, RA, Crohn's disease, sarcoidosis, leprosy, Castleman's disease, Takayasu's arteritis, multiple myeloma, and pristane-induced arthritis (163,185-186). Furthermore, IL-6 transgenic mice show a striking increase in agalactosyl IgG, suggesting that IL-6 may directly increase the level of agalactosyl IgG (185). Such evidence further strengthens the intimate relationship between IL-6 and certain autoimmune diseases and/or idiopathic diseases.(to The table of contents)
IL-6 transgenic mice develop a massive polyclonal plasmacytosis with autoantibodies and mesangial cell proliferative glomerulonephritis, resembling the autoimmune diseases observed in NZB/W F1 mice or SLE patients (45). The development of hypergammaglobulinemia has also been reported to occur in mice whose bone marrow has been altered by transplantation of cells infected with a retroviral vector expressing murine IL-6 (46). The involvement of IL-6 in autoimmune insulin dependent type I diabetes is suggested by IL-6 transgenic mice, which overexpress IL-6 in the pancreatic islet b cells (187). Although these transgenic mice remain normoglycemic through their lives, histopathological examination reveals that, particularly in older mice, there is a florid insulitis that is composed predominantly of B220+ B lymphocytes and to a lesser extent, Mac-1+ macrophage and T lymphocytes. Furthermore, the infiltration of plasma cells in the peri-islet is observed. The facts support the notion that the overexpression of IL-6 induces B-cell differentiation and may play roles in autoimmune disease. This concept is supported by the fact that anti-IL-6 antibodies inhibit the development of insulin-dependent diabetes in NOD/Wehi mice (188). Chronic administration of anti-IL-6 rat monoclonal antibodies together with anti-CD4, which induces tolerance to rat immunoglobulin, prevents the production of anti-dsDNA, reduces proteinuria, and prolongs life (189). These results suggest that IL-6 is one of critical factors in the pathogenesis of autoimmune diseases.
Plasma cells generated in IL-6 transgenic mice are not transplantable to syngeneic animals, indicating that for the malignant transformation, additional factors may be required (45). In this context, it is interesting to consider the fact that susceptibility to pristane-induced plasmacytomagenesis is genetically determined and that most inbred strains other than BALB/c are resistant (190). Consistent with this, the C57BL/6 IL-6 transgenic mice when backcrossed to BALB/c mice show a progression from polyclonal plasmacytosis to fully transformed monoclonal plasmacytoma, which displays chromosomal translocation with c-myc gene rearrangement (47). The evidence strongly supports the hypothesis that deregulated expression of the IL-6 gene can trigger polyclonal plasmacytosis resulting in the generation of malignant monoclonal plasmacytoma (48). Consistent with this notion, IL-6 deficient mice do not develop plasmacytoma (49), indicating the critical role of IL-6 in murine plasmacytomagenesis.(to The table of contents)
Both genetic and environmental factors play critical roles in autoimmune disease, although the degree to which each factor contributes to pathogenesis is different among autoimmune diseases. Several genetic factors including MHC, TNF, Fas, and Fas ligand have been proposed to be involved in autoimmune disease (191-194). Among them, MHC antigens play a critical role in selecting and/or activating autoreactive T-cell clones. On the other hand, several environmental factors, such as bacterial infection, viral infection, and continuous stimulation by foreign materials also play roles. In fact, pristane induces RA-like arthritis in certain strains of mice (165). Based on these facts, I have proposed a hypothesis explaining the molecular mechanisms that underlie certain autoimmune diseases, such as RA (195,196). Fig. 6 illustrates a hypothesis that is slightly modified from the one originally proposed. Furthermore, I have proposed a novel disease category termed, "chronic inflammatory proliferative disease," or CIPD (197). CIPD is characterized by the presence of an immune response, an inflammatory response, and the chronic proliferation of pathogenic cells accompanied by the constitutive expression of multiple genes. CIPD may include RA, mesangial proliferative glomerulonephritis, psoriasis, and artheriosclerosis. In CIPD, the continuous proliferation of pathogenic cells is observed, such as synovial cells in RA, mesangial cells in glomerulonephritis, and endotherial smooth muscle cells in arteriosclerosis. Certain autoimmune diseases may belong to CIPD.
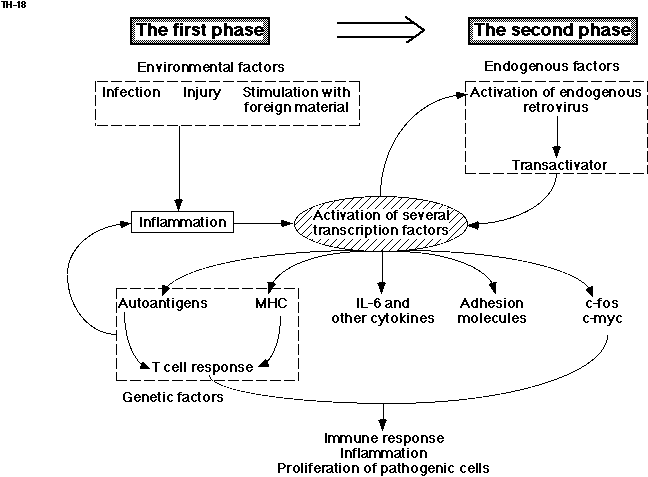
In the hypothesis illustrated in Fig. 6, the constitutive activation of a set of transcription factors, such as NF-kB, is the central factor governing the onset as well as the progression of CIPD. There are at least two phases in the course of CIPD. In the initial phase, the activation of the transcription factors is induced by a variety of stimuli, including infection, stimulation with foreign materials such as pristane, and injury. This initial stimulation induces the expression of a variety of cytokines that activate a set of transcription factors, leading to the expression of various genes encoding cellular proteins, including IL-6 and other cytokines, MHC molecules, adhesion molecules, various autoantigens such as heat shock proteins, and several transcription factors. Thus expressed, autoantigens could be recognized by autoreactive T-cells in the context of the MHC molecules. Such an autoreactive T-cell response is genetically determined but could also be enhanced by IL-6 and other cytokines. In certain autoimmune diseases that are more dependent on genetic factors, the autoreactive T-cell activation must be the primary response. Thus activated autoreactive T-cells also induce the inflammatory response. IL-6 can cause polyclonal B-cell activation and the auto-reactive B-cell response. The enhanced expression of several adhesion molecules on bone marrow stromal cells also induce an increase in pre-B-cell growth and differentiation (198,199). Several transcription factors and cytokines further modulate the expression of a variety of cellular genes that affect inflammatory reactions and cell growth, leading to the proliferation of pathogenic cells.
The first phase could be followed by the second phase, in which the activation of a set of transcription factors is under the control of endogenous factorsハ in certain autoimmune diseases, in particular the diseases categorized as CIPD. For such endogenous factors, I hypothesize the involvement of a transactivator derived from an endogenous retrovirus. During the first phase, certain activated transcription factors, such as NF-kB, could activate the resting endogenous retrovirus. This activated retrovirus could produce a transactivator, such as p40tax, that could further activate a set of cellular transcription factors. Once such an activation loop is turned on, multiple genes are continuously expressed independently of normal regulatory mechanisms. Therefore, even if the initial stimuli are withdrawn, the constitutive expression of multiple genes could be continued in the second phase. In this way, the molecular mechanism of CIPD and some autoimmune diseases, in particular RA, could be explained. Evidence has been accumulated suggesting the involvement of viruses in human autoimmune diseases (200-202). In fact, HTLV-1 transgenic mice develop inflammatory arthropathy resembling RA (203). Furthermore, virus-derived transactivator, p40tax, and HIV tat have been shown to actually induce a variety of cytokines including IL-6, or their receptor gene expression (204-211). Interestingly, pristane, which induces RA-like arthritis in mice, activates an endogenous retrovirus (212). Furthermore, it is suggested that the constitutive production of IL-6 by fibroblasts that are derived from patients with systemic sclerosis, is mediated by the constitutive binding of nuclear factors to the IL-6 promoter (213). Therefore, future studies on the molecular mechanism(s) of the dysregulated expression of the IL-6 gene or other genes in patients will clarify the pathogenesis of autoimmune diseases and CIPD.(to The table of contents)
Conclusion and future prospects
Studies of IL-6 and its receptor performed during the last decade have revealed much about the molecular basis of cytokine biology. IL-6 is a typical cytokine and studies on the IL-6 system have greatly contributed to the establishment of many general concepts about the cytokine system. The molecular mechanisms regulating cell growth and differentiation through cytokine receptors have been elucidated. We have learned much about the roles of IL-6 in vivo from studies using IL-6 transgenic mice and IL-6 deficient mice. Furthermore, it has been demonstrated that IL-6 and other inflammatory cytokines play critical roles in the formation of the pathological states of a variety of diseases, in particular, chronic inflammation, autoimmune diseases, viral infection, and malignancies. IL-6 may be critically involved in the cause of certain diseases, such as myeloma, plasmacytoma and postmenopausal osteoporosis.
We now know much about IL-6, its receptor, and the signal-transduction mechanisms of IL-6 and other cytokines. However, it is also true that there is much we still do not know about them. Is there any yet unidentified novel cytokine exerting important biological functions? Is there any yet unidentified novel signal-transduction pathway ? How do multiple signal-transduction pathways through a cytokine receptor interact with each other, and how do they eventually regulate and determine the cell fate: growth and differentiation? Is it possible for us to control the action of cytokines completely ? If so, how can we do it ? During the next decade, we will know much more, about the mode of interaction between cytokines and their receptors, the molecular mechanisms of many signal transducing molecules, and the in vivo roles of these molecules in both health and disease. These studies will provide the methods and materials to regulate the action of cytokines. Ultimately, it will be possible for us to control the immune response, inflammation, and hematopoiesis in both healthy and diseased states.(to The table of contents)
Acknowledgments
I thank many colleagues, in particular, Drs. Koichi Nakajima, Masahiko Hibi, Yoshio Fujitani, Yojiro Yamanaka, Toshiyuki Fukada, and Mariko Takahashi-Tezuka, who contributed to our recent studies described in this review. I also thank Ms. R. Masuda and T. Kimura for their excellent secretarial assistance. This study was supported in part by Grant-in-Aid for COE Research of the Ministry of Education, Science, Sports and Culture in Japan.
1. Hirano, T., Yasukawa, K., Harada, H., Taga, T., Watanabe, Y., Matsud, T., Kashiwamura, S., Nakajima, K., Koyama, K., Iwamatsu, A., Tsunasawa, S., Sakiyama, F., Matsui, H., Takahara, Y., Taniguchi, T., and Kishimoto, T. Complementary DNA for a novel human inteleukin (BSF-2) that induces B lymphocytes to produce immunoglobulin. Nature 324, 73-76,1986.(Abstract)
2. Zilberstein, A., Ruggieri, R., Korn, J. H., and Revel, M. Structure and expression of cDNA and genes for human interferon-beta-2, a distinct species inducible by growth-stimulatory cytokines. EMBO J. 5, 2529-2537, 1986.
3. May, L. T., Helfgott, D. C., and Sehgal, P. B., Anti-b-interferon antibodies inhibit the increased expression of HLA-B7 mRNA in tumor necrosis factor-treated human fibroblasts: Structural studies of the b2 interferon involved. Proc Natl Acad Sci USA 83, 8957-8961, 1986.
4. Haegeman, G., Content, J., Volckaert, G., Derynck, R., Tavernier, J., and Fiers, W. Structural analysis of the sequence encoding for an inducible 26-kDa protein in human fibroblasts. Eur J Biochem 159, 625-632, 1986.
5. Muraguchi, A., Kishimoto, T., Miki, Y., Kuritani, T., Kaieda, T., Yoshizaki, K., and Yamamura,Y. T cell-replacing factor ( TRF )-induced IgG secretion in human B blastoid cell line and demonstration of acceptors for TRF. J Immunol 127, 412-416, 1981.
6. Teranishi, T., Hirano, T., Arima, N., and Onoue, K. Human helper T cell factor(s) (ThF). II. Induction of IgG production in B lymphoblastoid cell lines and identification of T cell replacing factor (TRF)-like factor(s). J. Immunol 128, 1903-1908, 1982.(Abstract)
7. Okada, M., Sakaguchi, N., Yoshimura, N., Hara, H., Shimizu, K., Yoshida, N., Yoshizaki, K., Kishimoto, S., Yamamura, Y., and Kishimoto, T. B cell growth factor (BCGF) and B cell differentiation factor from human T hybridomas: Two distinct kinds of BCGFs and their synergism in B cell proliferation. J. Exp Med 157, 583-590, 1983.
8. Hirano, T., Teranishi, T., and Onoue, K. Human helper T cell factor(s) III. Characterization of B cell differentiation factor I (BCDF I). J. Immunol. 132, 229-234, 1984.(Abstract)
9. Hirano, T., Teranishi, T., Lin, B. H., and Onoue, K. Human helper T cell factor(s) IV. Demonstration of a human late-acting B cell differentiation factor acting on Staphylococcus aureus Cowan I-stimulated B cells. J. Immunol. 133,798-802, 1984.(Abstract)
10. Hirano, T., Taga, T., Nakano, N., Yasukawa, K., Kashiwamura, S., Shimizu, K., Nakajima, K., Pyun, K. H., and Kishimoto, T. Purification to homogeneity and characterization of human B cell differentiation factor (BCDF or BSFp-2). Proc. Natl. Acad. Sci., USA., 82, 5490-5494, 1985.(Abstract)
11. Sehgal, P. B., May, T. T., Tamm, I., and Vilcek, J. Human b2 interferon and B-cell differentiation factor BSF-2 are identical. Science 235, 731-732, 1987.
12. Hirano, T., and Kishimoto ,T., Interleukin 6. In: Handbook of Experimental Pharmacology. vol. 95/I , Peptide growth factors and their receptors I 1990, edited by Sporn, M. B. and Roberts, A. B. Berlin: Springer-Verlag, pp. 633-665.
13. Van Damme, J., Opdenakker, G., Simpson, R. J. Rubira, M. R., Cayphas, S., Vink, A., Billiau, A., and Snick, J. V. Dentification of the human 26-kDa protein, interferon b2 (IFNb2), as a B cell hybridoma/plasmacytoma growth factor induced by interleukin 1 and tumor necrosis factor. J Exp Med 165, 914-919, 1987.
14. Van Snick, J., Cayphas, S., Szikora, J. P., Renauld , J. C., Van Roost, E., Boon , T., Simpson, R. J. cDNA cloning of murine interleukin-HP1:homology with human interleukin 6. Eur J. Immunol 18, 193-197, 1988.
15. Aarden, L., Lansdorp, P., De Groot, E., A growth factor for B cell hybridomas produced by human monocytes. Lymphokines 10, 175-185, 1985.
16. Nordan, R. P., and Potter, M. A macrophage-derived factor required by plasmacytomas for survival and proliferation in vitro. Science 233, 566-569, 1986.
17. Kawano, M., Hirano, T., Matsuda, T., Taga, T., Horii, Y., Iwato, K., Asaoku, H., Tang, B., Tanabe, O., Tanaka, H., Kuramoto, A., and Kishimoto, T. Autocrine generation and essential requirement of BSF-2/IL-6 for human multiple myelomas. Nature , 332, 83-85, 1988.(Abstract)
18. Gauldie, J., Richards, C., Harnish, D., Lansdorp, P., and Baumann, H. Interferon b2/B-cell stimulatory factor type 2 shares identity with monocyte-derived hepatocyte-stimulating factor and regulates the major acute phase protein response in liver cells. Proc Natl Acad Sci USA 84, 7251-7255, 1987.
19. Andus, T., Geiger, T., Hirano, T., Northoff , H., Ganter, U., Bauer, J., Kishimoto, T., Heinrich, P. C., Recombinant human B cell stimulatory factor 2 (BSF-2/IFNァ2) regulates ァ-fibrinogen and albumin mRNA levels in Fao-9 cells. FEBS Lett 221, 18-22, 1987.
20. Sehgal, P. B., Grienger, G., and Tosato, G. (editors) Regulation of the Acute Phase and Immune Responses: Interleukin-6. Ann New York Acad Sci 557, 1-583, 1989.
21. Van Snick, J. Interleukin-6: An overreviw. Annu Rev Immunol 8, 253-278, 1990.
22. Heinrich, P. C., Castell, J. V., and Andus, T. Interleukin-6 and the acute phase response. Biochem J. 265, 621-636, 1990.
23. Hirano, T. The biology of interleukin 6. Chemical Immunol 51, 153-180, 1992.
24. Hirano, T. Interleukin 6. In: The Cytokine Handbook, 2nd edition 1994, edited by Thomson, A. W. London: Academic Press, pp145-168, 1994.
25. Bazan, J. F. Haemopoietic receptors and helical cytokines. Immunol Today 11, 350-354, 1990.
26. Bazan, J. F. Neurotropic cytokines in the hematopoietic fold. Neuron 7, 1-12, 1992.
27. Bazan, J. F. A novel family of growth factor receptors: A common binding domain in the growth hormone, prolactin, erythropoietin and IL-6 receptors, and the p75 IL-6 receptor-chain. Biochem. Biophys. Res. Comm. 164, 788-795, 1989.
28. Bazan, J. F. Structural design and molecular evolution of a cytokine receptor superfamily. Proc. Natl. Acad. Sci. USA 87, 6934-6938, 1990.
29. Miyajima, A., Kitamura, T., Harada, N., Yokota, T., and Arai, K. Cytokine receptors and signal transduction. Annu. Rev. Immunol. 10, 295-339, 1992.
30. Sugamura, K., Asao, H., Kondo, M., Tanaka, N., Ishii, N., Nakamura, M., and Takeshita, T. The common gamma-chain for multiple cytokine receptors. Adv Immunol 59, 225-277,1995.
31. Taniguchi, T. Cytokine signaling through nonreceptor protein tyrosine kinase. Science 268, 251-255, 1995.
32. Kishimoto, T., Akira, S., Narazaki, M., and Taga, T. Interleukin-6 family of cytokines and gp130. Blood, 86, 1243-1254, 1995.
33. Hibi, M., Nakajima, K., and Hirano, T. IL-6 cytokine family and signal transduction: a model of the cytokine system. J. Mol. Med. 74, 1-12, 1996.(Abstract)
34. Taga, T., Hibi, M., Hirata, Y., Yamasaki, K., Yasukawa, K., Matsuda, T., Hirano, T., and Kishimoto, T. Interleukin 6 (IL-6) triggers the association of its receptor (IL-6-R) with a possible signal transducer, gp130. Cell 58, 573-581, 1989.(Abstract)
35. Hibi, M., Murakami, M., Saito, M., Hirano, T., Taga, T., and Kishimoto, T. Molecular cloning and expression of an IL-6 signal transducer, gp130. Cell 63, 1149-1157, 1990.(Abstract)
36. Gearing, D. P., Comeau, M. R., Friend, D. J., Gimpel Thut, C. J., Mcgourty, J., Brasher, K. K., King, J. A., Gills, S., Mosley, B., Ziagler, S. F., and Cosman, D. The IL-6 signal transducer, gp130: an oncostatin M receptor and affinity converter for the LIF receptor. Science 255, 1434-1437, 1992.
37. Ip, N.Y., Nye, S. H., Boulton, T. G. , Davis, S., Taga, T., Li, Y., Birren, S. J., Yasukawa, K., Kishimoto, T., Anderson, D. J., Stahl, N., and Yancopoulos, G. D. CNTF and LIF act on neuronal cells via shared signaling pathways that involve the IL-6 signal transducing receptor component gp130. Cell 69, 1121-1132, 1992.
38. Yin, T., Taga, T., Tsang, M L.-S, Yasukawa, K., Kishimoto, T., and Yang, Y.-C Involvement of IL-6 signal transducer gp130 in IL-11-mediated signal transduction. J. Immunol. 151, 2555-2561, 1993.
39. Taga, T., Narazaki, M., Yasukawa, K., Saito, T., Miki. D., Hamaguchi, M., Davis, S., Shoyab, M., Yancopoulos, G. D., and Kishimoto, T. Functional inhibition of hematopoietic and neurotrophic cytokines by blocking the interleukin 6 signal transducer gp130. Proc. Natl. Acad. Sci. USA 89, 10998-11001, 1992.
40. Pennica, D., King, K. L., Shaw, K. L., Luis, E., Rullamas, J., Luoh, S., Darbonne, W. C., Knutzon, D. S., Yen, R., Chien, K. R., Baker, J. B., Wood, W. I. Expression cloning of cardiotrophin-1, a cytokine that induces cardiac myocyte hypertrophy. Proc. Natl. Acad. Sci. USA 92, 1142-1146, 1995.
41. Ihle, J. N., Witthuhn, B. A., Quelle, F. W., Yamamoto, K., Thierfelder, W. E., Kreider, B., Silvennovinen, O. Signaling by the cytokine receptor superfamily, JAKs and STATs. TIBS 19, 222-227, 1994.
42. Darnell, J. E., Kerr, I. M., Stark, G. M., Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 257, 803-806, 1994.
43. Schindler, C., and Darnell, J. E., Jr. Transcriptional responses to polypeptide ligands:The JAK-STAT pathway. Annu. Rev. Biochem. 64, 621-651, 1995.
44. Ihle, J. N. STATs: Signal transducers and Activators of Transcription. Cell 84, 331-334,1996.
45. Suematsu, S., Matsuda, T., Aozasa, K., Akira, S., Nakano, N., Ohno, S., Miyazaki, J., Yamamura, K., Hirano, T., and Kishimoto, T. IgG1 plasmacytosis in interleukin 6 transgenic mice. Proc. Natl. Acad. Sci. USA 86, 7547-7551, 1989.(Abstract)
46. Brandt, S. J., Bodine, D. M., Dunbar, C. E., Nienhuis, A. Dysregulated interleukin 6 expression produces a syndrome resembling Castleman's disease in mice. J. Clin Invest 86, 592-599, 1990.
47. Suematsu, S., Matsusaka, T., Matsuda, T., Ohno, S., Miyazaki, J., Yamamura, K., Hirano, T., and Kishimoto, T. Generation of transplantable plasmacytomas with t(12;15) in Il-6 transgenic mice. Proc. Natl. Acad. Sci. USA 89, 232-235, 1992.(Abstract)
48. Hirano T: Interleukin 6 (IL-6) and its receptor: Their role in plasma cell neoplasias. Int J Cell Cloning 9, 166-184, 1991.
49. Hilbert, D. M., Kopf, M., Mock, B. A., Kohler, G., and Rudikoff, S. Interleukin 6 is essential for in vivo development of B lineage neoplasms. J. Exp. Med. 182, 243-248, 1995.
50. Kopf, M., Baumann, H., Freer, G., Freudenberg, M., Lamers, M., Kishimoto, T., Zinkernagel, R., Bluethmann, H., and Kohler, G. Impaired immune and acute-phase responses in interleukin-6-deficient mice. Nature 368, 339-342, 1994.
51. Beagley, K.W., Eldridge, J. H., Lee, F., Kiyono ,H., Everson, M. P., Koopman, W. J., Hirano, T., Kishimoto, T., and McGhee, J. R. Interleukins and IgA synthesis: Human and murine IL-6 induce high rate IgA secretion in IgA-committed B cells. J. Exp. Med. 169, 2133-2148, 1989.
52. Fujihashi, K., McGhee, J. R., Lue, C., Beagley, K. W. Taga, T., Hirano, T. Kishimoto, T. Mestecky, J. and Kiyono, H. . Human appendix B cells naturally express receptors for and respond to interleukin 6 with selective IgA1 and IgA2 synthesis. J. Clin. Invest 88, 248-52,1991.
53. Ramsay, A. J., Husband, A. J., Ramshaw, I. a., Bao, S., Matthaei, K. I., Koehler G., and Kopf, M. The role of interleukin-6 in mucosal IgA antibody responses in vivo. Science 264, 561-563, 1994.
54. Dalrymple, S. A., Lucian, L. A., Slattery, R., McNeil, T., Aud, D. M., Fuchino, S., Lee, F., and Murray, R. Interleukin-6-deficient mice are highly susceptible to Listeria monocytogenes infection: correlation with inefficient neutorphilia. Infect. Immun. 63, 2262-2268, 1995.
55. Jilka, R. L., Hangoc, G., Girasole, G., Passeri, G., Williams, D. C., Abrams, J. S., Boyce B, Broxmeyer H, Manolagas SC() Increased osteoclast development after estrogen loss: mediation by interleukin-6. Science 257, 88-91, 1992.
56. Girasole, G., Jilka, R. L., Passeri, G., Boswell, S., Boder, G., williams, D. C. and Manolagas, S. C. 17 beta-estradiol inhibits interleukin-6 production by bone marrow-derived stromal cells and osteoblasts in vitro: a potential mechanism for the antiosteoporotic effect of estrogens. J. Clin. Invest. 89, 883-891, 1992.
57. Poli, V., Balena, R., Fattori, E., Markatos, a., Yamamoto, M., Tanaka, H., Ciliberto, G., Rodan, G. A., and Costantini, F. Interleukin-6 deficient mice are protected from bone loss caused by estrogen depletion. EMBO J. 13, 1189-1196, 1994.
58. Chai, Z., Gatti, S., Toniatti, C., Poli, V., and Barfai, T. Interleukin(IL)-6 gene expression in the central nervous system is necessary for fever response to lipopolysaccharide or IL-1 beta: a study on IL-6-deficient mice. J. Exp. Med. 183, 311-316, 1996.
59. Gearing, D., Gough, N. M., King, J. A., Hilto, D. J., Nicola, N. A., Simpson, R. J., Nice, E. C., Kelso, A., Metcalf, D., Molecular cloning and expression of cDNA encoding a murine myeloid leukaemia inhibitory factor (LIF). EMBO J. 6, 3995-4002, 1987.
60. Hilton, D. J. LIF: lots of interesting functions. Trends Biochem. Sci. 17, 72-76, 1992.
61. Williams, R. L., Hilton, D. J., Pease, S., Wilson, T. A., Stewart, C. L., Gearing, D. P., Wagner, E. F., Metcalf, D., Nicola, N. A., Gough, N. M. Myeloid leukemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature 336, 684-687, 1988.
62. Mori, M., Yamaguchi, K., and Abe, K. Purification of a lipoprotein lipase-inhibiting protein produced by a melanoma cell line associated with cancer cachexia. Biochem. Biophys. Res. Commun. 160, 1085-1092, 1989.
63. Yamamori, T., Kukada, K., Abersold, R., Korsching, S., Fann, M. J., and Patterson, P. H. The cholinergic neuronal differentiation factor from heart cells is identical to leukemia inhibitory factor. Science 246, 1412-1416, 1989.
64. Lin, L.-F. H., Mismer, D., Lile, J. D., Armes, L. G., Butler, E. T. III, Vannice, J. L., and Collins, F. Purification, cloning, and expression of ciliary neurotrophic factor (CNTF). Science 246, 1023-1025, 1989.
65. Adler, R., Landa, K., Manthorpe, M., and Varon, S. Cholinergic neurotrophic factors: Intraocular distribution of trophic activity for ciliary neurons. Science 204, 1434-1436, 1979.
66. Barbin, G., Manthorpe, M., and Varon, S. Purification of chick eye ciliary neurotrophic factor. J. Neurochem. 43, 1468-1478, 1984.
67. Manthorpe, M., Skaper, S., Williams, L. R., and Varon, S. Purification of adult rat sciatic nerve ciliary neurotrophic factor. Brain. Res. 367, 282-286, 1986.
68. Ernsberger, U., Sendtner, M., and Rohrer, H. Proliferation and differentiation of embryonic chick symphathetic neurons: effects of ciliary neurotrophic factor. Neuron 2, 1275-1284, 1989.
69. Saadat, S., Sendtner, M., Rohrer, H. Ciliary neurotrophic factor induces cholinergic differentiation of rat sympathetic neurons in culture. J. Cell. Biol. 108, 1807-1816, 1989.
70. Hughes, S. M., Lillien, L. E., Raff, M. C., Rohrer, H., and Sedtner, M. Ciliary neurotrophic factor induces type-2 astrocyte differentiation in culture. Nature 335, 70-73, 1988.
71. Sendtner, M., Kreutzberg, G. W., and Thoenen, H. Ciliary neurotrophic factor prevents the degeneration of motor neurons after axotomy. Nature 345, 440-441, 1990.
72. Murphy, M., Reid, K., Hilton, D.J., and Bartlett, P. F. Generation of sensory neurons is stimulated by leukemia inhibitory factor. Proc. Natl. Acad. Sci. USA 88,3498-3501, 1991.
73. Malik, N., Kalestad, J. C., Gunderson, N. L., Austin, S. D., Neubauer, M. G., Ochs, V., Marquardt, H., Zarling, J. M., Shoyab, M., Wei, C.-M., Linsley, P., Rose, T. M. Molecular cloning, sequence analysis, and functional expression of a novel growth regulator, oncostatin M. Mol. Cell. Biol. 9, 2847-2853, 1989.
74. Zarling, J. M., Shoyab, M., Marquardt, H., Hanson, M. B., Lioubin, M. N., and Todaro, G. J. Oncostatin M: A growth regulator produced by differentiated histiocytic lymphoma cells. Proc. Natl. Acad. Sci. USA 83, 9739-9743, 1986.
75. Nair, B. C., DeVico, A. L., Nakamura, S., Copeland, T. D., Chen, Y., Patel, A., O'Neil, T., Oroszlan, S., Gallo, R. C., and Sarngadharan, M. G. Identification of a major growth factor for AIDS-Kaposi's sarcoma cells as oncostatin M. Science 255, 1430-1432, 1992.
76. Miles, S. A., Martinez-Maza, O., Rezai, A., Magpantay, L., Kishimoto, T., Nakamura, S., Radka, S. F., and Linsley, P. S. Oncostatin M as a potent mitogen for AIDS-Kaposi's sarcoma-derived cells. Science 255, 1432-1434, 1992.
77. Paul, S. R., Bennett, F., Calvetti, J. A., Kelleher, K. , Wood, C. R., O'Hara, Jr. R. M., Leary, A. C., Sibley, B. , Clark, S. C., Williams, D. A., and Yang, Y.-C. Proc. Natl. Acd. Sci. USA 87, 7512-7516, 1990.
78. Yin, T., Schendel, P., Yang, Y. C. Enhancement of in vivo and in vitro antigen-specific antibody responses by interleukin-11. J. Exp. Med. 175, 211-216, 1992.
79. Pennica, D., Shaw, T. A., Swanson, M. W., Moore, D. L., Sjelton, K. A., Zioncheck, A., Rosenthal, A., Taga, T., Paoni, N. F., and Wood, W. I. Cardiotrophin-1. Biological activities and binding to the leukemia inhibiory factor receptor/gp130 signaling complex. J. Biol. Chem. 270, 10915-10922, 1995.
80. Metcalf, D. Actions and interactions of G-CSF, LIF, and IL-6 on normal and leukemia murine cells. Leukemia 3, 349-355, 1989.
81. Rose, T. M., Bruce, A. G. Oncostatin M is a member of a cytokine family which includes leukemia inhibitory factor, granulocyte colony-stimulatory factor and interleukin-6. Proc. Natl. Acad. Sci. USA 88, 8641-8645, 1991.
82. Zhang, X. G., Gu, J. J, Lu, Z. Y., Yasukawa, K., Yancopoulos, G. D., Turner, K., Shoyab, M., Taga, T., Kishimoto, T., Bataille, R., and Klein, B. Ciliary neurotrophic factor, interleukin 11, leukemia inhibitory factor, and oncostatin M are growth factors for human myeloma cell lines using the interleukin 6 signal transducer gp130. J. Exp. Med. 179, 1337-1342, 1994.
83. Musashi, M., Yang, Y. C., Paul, S. R., Clark, S. C., Sudo, T., Ogawa, M. Direct and synergistic effects of interleukin-11 on murine hemopoiesis in culture. Proc. Natl. Acad .Sci. USA, 88, 765-769, 1991.
84. Leary, A. G., Zeng, H. Q., Clark, S. C., and Ogawa, M. Growth factor requerements for survival in G0 and entry into the cell cycle of primitive human hemopoietic progenitors. Proc. Natl. Acd. Sci. USA 89, 4013-4017, 1992.
85. Baumann, H., Onorato, V., Gauldie, J., and Jahreis, G. P. Distinct sets of acute phase plasma proteins are stimulated by separate human hepatocyte-stimulating factors and monokines in rat hepatoma cells. J. Biol. Chem. 262, 9756-9768, 1987.
86. Baumann, H., and Wong, G. G. Hepatocyte-stimulatory factor III shares structural and functional identity with leukemia inhibitory factor. J. Immunol. 143, 1163-1167, 1989.
87. Richards, C.D., Brown, T. J., Shoyab, M., Baumann, and H., Gauldie, J. Recombinant oncostatin M stimulates the production of acute phase protein in HepG2 cells and rat primary hepatocytes in vitro. J. Immunol. 148, 1731-1736, 1992.
88. Baumann, H., and Schendel, P. Interleukin 11 regulates the hepatic expression of the same plasma protein genes as interleukin-6. J. Biol. Chem. 266, 20424-20427, 1991.
89. Patterson, P. H., and Nawa, H. Neuronal differentiation factors/cytokines and synaptic plasticity. Cell 72, 123-137, 1993.
90. Yamasaki, K., Taga, T., Hirata, Y., Yawata, H., Kawanishi, Y., Seed, B., Taniguchi, T,, Hirano. T., and Kishimoto, T. Cloning and expression of the human interleukin-6 ( BSF-2/IFNb2 ) receptor. Science 241, 825-828, 1988.
91. Sugita, T., Totsuka, T., Saito, M., Yamasaki. K., Taga, T., Hirano, T., and Kishimoto, T. Functional murine IL-6 receptor with the intracisternal A-particle gene product at its cytoplasmic domain: Its possible role in plasmacytomagenesis. J. Exp. Med. 171, 2001-2009, 1990.
92. Paonessa, G., Graziani, R., Serio, A. D., Svio, R., Ciappori, L., Lahm, A., Salvati, A. L., Tniatti, C., and Ciliberto, G. Two distinct and independent sites on IL-6 trigger gp130 dimer formation and signalling. EMBO J. 14, 1942-1951, 1995.
93. Kobayashi, M., Fitz, L., Ryan, M., Hewick, R. M., Clark, S. C., Chan, S., Loudon, R., Sherman, F., Perrussia, B., and Trinchieri, G. Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J. Exp. Med. 170, 827-845, 1989.
94. Gearing, D. P., and Cosman, D. Homology of the p40 subuniot of natural killer cell stimulatory factor (NKSF) with the extracellular domain of the interleukin-6 receptor. Cell 66, 9-10, 1991.
95. Chua, A. O., Chizzonite, R., Desai, B. B., Truitt, T. P., Nunes, P., Minetti, L. J., Warrier, R. R., Presky, D. H., Levine J. F., Gately. M. K., and Gubler, U. Expression cloning of a human IL-12 receptor component. A new member of the cytokine receptor superfamily with strong homology to gp120. J. Immunolo. 153, 128-136, 1994.
96. Davis, S., Aldrich, T. H., Ip, N. Y., Stahl, N., Scherer, S., Farruggella, T., DiStefano, P. S., Curtis, R., Panayotatos, N., Gascan, H., Chevalier, S., and Yancopoulos, G. D. Released form of CNTF receptor a components as a soluble mediator of CNTF responses. Science 259, 1736-1739, 1993.
97. Hirota, H., Yoshida, K., Kishimoto, T., and Taga, T. Continuous activation of gp130, a signal transducing receptor component for interleukin 6-related cytokines, causes myocardial hypertrophy in mice. Proc. Natl. Acad. Sci. USA. 92, 4862-4866, 1995.
98. Treanor, J. J. S., Goodman, L., de Sauvage, F., Stone, D. M., Poulsen, K. T., Beck, C. D., Gray, C., Armanini, M. P., Pollock R. A., Hefti, F., Phillips H. S., Goddard, A., Moore, M. W., Buj-Bello, A., Davies, A. M., Asai, N., Takahashi, M., Vandlen R., Henderson, C. E., and Rosenthal, A. Characterization of a multicomponent receptor for GDNF. Nature, 382, 80-83, 1996.
99. Jing, S., Wen, D., Yu, Y., Holst P. L., Luo, Y., Fang, M., Tamir, R., Antonio, L., Hu, Z., Cupples, R., Louis, J.-C., Hu, S., Altrock, W. B., and Fox, G. M. GDNF-Induced Activation of the Ret Protein Tyrosine Kinase Is Mediated by GDNFR-a, a Novel Receptor for GDNF. Cell 85, 1113-1124, 1996.
100. Velazquez, L., Fellous, M., Stark, G. R., and Pellegrini, S. A protein tyrosine kinase in the interferon a/b signaling pathway. Cell 70, 313-322, 1992.
101. Witthuhn, B. A., Quelle, F. W., Silvennoinen, O., Yi, T., Tang, B,. Miura, O., and Ihle, J. N. Jak2 associates with the erythropoietin receptor and is tyrosine phosphorylated and activated following stimulation with erythropoietin. Cell 74, 227-236, 1993.
102. Argetsinger, L. S., Campbell, G. S., Yang, X., Witthuhn, B. A., Silvennoinen, O., Ihle, J. N., and Carter,-Su. C. Identification of Jak2 as a growth hormone receptor-associated tyrosine kinase. Cell 74, 237-244, 1993.
103. Lutticken, C., Wegenka, U. M., Yuan, J., Buschmann, J., Schindler, C., Ziemiecki, A., Harpur, A. G., Wilks, A. F., Yasukawa, K., Taga, T., Kishimoto, T., Barbieri, G., Pellegrini, S., Sendtner, M., Heinrich, P. C., and Horn, F. Association of transcription factor APRF and protein kinase Jak1 with the interleukin-6 signal transducer gp130. Science 263, 89-92, 1993.
104. Stahl, N., Boulton, T. G., Farruggella, T., Ip, N. Y., Davis, S., Witthuhn, B. A., Quelle, F. W., silvernnoinen, O., Barbieri, G., Pellgrini, S., Ihle, J. N., and Yancopoulos, G. D. Association and activation of Jak/Tyk kinases by CNTF/LIF/OSM/IL-6 b receptor components. Science 263, 92-95, 1993.
105. Matsuda, T., Yamanaka, Y., and Hirano, T. Interleukin-6-induced tyrosine phophorylation of multiple proteins in murine hematopoietic lineage cells. Biochem. Boiphys. Res. Comm. 200, 821-828, 1994.
106. Akira, S., Nishio, Y., Inoue, M., Wang, X.J., Wei, S., Matsusaka, T., Yoshida, K., Sudo, T., Naruto, M., and Kishimoto, T. Molecular cloning of APRF, a novel IFN-stimulated gene factor3 p91-related transcriptional factor involved in the gp130-mediated signaling pathway. Cell 77, 63-71, 1994.
107. Zhong, Z., Wen, Z., and Darnell, J. E. Stat3: a STAT family member activated by tyrosine phosphorylation in response to Epidermal Growth Factor and interleukin-6. Science 264, 95-98, 1994.
108. Fujitani, Y., Nakajima, K., Kojima, H., Nakae, K., Takeda, T., and Hirano, T. Transcriptional activation of the IL-6 responsive element in the junB promoter is mediated by multple Stat family proteins. Biochem. Biophys. Res. Commun. 202, 1181-1187, 1994.(Abstract)
109. Nakajima, K., Matsuda, T., Fujitani, Y., Kojima, H., Yamanaka, Y., Nakae, K., Takeda, T., and Hirano, T. Signal transduction through IL-6 receptor: Involvement of multiple protein kinases, Stat factors, and a novel H7-sensitive pathway. Annal of New York Academy of Science 762, 55-70, 1995.
110. Lai, C. F., J. Ripperger, K. K. Morella, Y. Wang, D. P. Gearing, N. D. Horseman, S. P. Campos, G. H. Fey, and H. Baumann. STAT3 and STAT5B are targets of two different signal pathways activated by hematopoietin receptors and control transcription via separate cytokine response elements. J. Biol.. Chem.. 270, 23254-23257, 1995.
111. Fujitani, Y., Hibi, M., Fukada, T., Takahashi-Tezuka, M., Yoshida, H., T. amaguchi, Y., Sugiyama, K., Yamanaka, Y., Nakajima, K., and Hirano, T. An alternative pathway for STAT activation that is mediated by the direct interaction between JAK and STAT. Oncogene. 14, 751-761, 1997. (Abstract)
112. Akira, S., H. Isshiki, T. Sugita, O. Tanabe, S. Kinoshita, Y. Nishio, T. Nakajima, T. Hirano, and T. Kishimoto. A nuclear factor for IL-6 expression(NF-IL-6) is a member of a C/EBP family. EMBO J. 9, 1897-1906, 1990.
113. Baumann, H., Morella, K. K., Campos, S. P., Cao, Z., and Jahreis, G. P. Role of CAAT-enhancer binding protein isoforms in the cytokine regulation of acute-phase plasma protein genes. J. Biol. Chem. 267, 19744-19751, 1992.
114. Baumann, H., Ziegler, S. F., Mosley, B., Morella, K. K., Pajovie, S., and Gearing, D. P. Reconstitution of the response to leukemia inhibitory factor, oncostatin M, and ciliary neurotrophic factor in hepatoma cells. J. BIol. Chem. 268, 8414-8417, 1993.
115. Wegenka, U. M., Buschmann, J., Lutticken, C., Heinrich, P. C., and Horn, F. Acute-phase response factor, a nuclear factor binding to acute-phase response elements, is rapidly activated by interleukin-6 at the posttranslational level. Mol. Cell. Biol. 13, 276-288, 1993.
116. Nakajima, K., Kusafuka, T., Takeda, T., Fujitani, Y., Nakae, K., and Hirano, T. Identification of a novel interleukin 6 responsive element containing an Ets-binding site and a CRE-like site in the junB promoter. Mol. Cell. Biol. 13, 3027-3041, 1993.(Abstract)
117) Kojima, H., Nakajima, K., and Hirano, T. IL-6-induced complexes on an IL-6 response element of the junB promoter contain Stat3 and 36kDa CRE-like site binding protein(s). Oncogene 12, 547-554, 1996.(Abstract)
118. Zhang, X., Blenis, J., Li, H. C., Schindler, C., and Chen,-Kiang. S. Requirement of serine phosphorylation for formation of STAT-promoter complexes. Science 267, 1990-1994, 1995.
119. Wen, Z., Zhong, Z., and Darnell, J. E. Maximal activation of transcription by Stat1 and Stat3 requires both tyrosine and serine phosphorylation. Cell 82, 241-250, 1995.
120. Fukunaga, R., Ishizaka-Ikeda, E., Pan, C. X., Seto, Y., and Nagata, S. Functional domains of the granulocyte colony-stimulating factor receptor. EMBO J. 10, 2855-2865, 1991.
121. Murakami, M., Narazaki, M., Hibi, M., Yawata, H., Yasukawa, K., Hamaguchi, M., Taga, T., and Kishimoto, T. Critical cytoplasmic region of the interleukin 6 signal transducer gp130 is conserved in the cytokine receptor family. Proc. Natl. Acad. Sci. U S A 88, 11349-11353, 1991.
122. Narazaki, M., B. A. Witthuhn, K. Yoshida, O. Silvennoinen, K. Yasukawa, J. N. Ihle, T. Kishimoto, and T. Taga. Activation of JAK2 kinase mediated by the interleukin 6 signal transducer gp130. Proc. Natl.. Acad.. Sci.. 91, 2285-2289, 1994.
123. Stahl, N., T. J. Farruggella, T. G. Boulton, Z. Zhong, J. J. Darnell, and G. D. Yancopoulos. Choice of STATs and other substrates specified by modular tyrosine- based motifs in cytokine receptors. Science 267, 1349-1353, 1995.
124. Yamanaka, Y., Nakajima, K., Fukada, T., Hibi, M., and Hirano, T. Differentiation and growth arrest signals generate through the cytoplasmic region of gp130 that is essential for Stat3 activation. EMBO J. 15, 1557-1565, 1996.(Abstract)
125. Baumann, H., Symes, A, J., Comeau, M. R., Morella, K. K., Wang, Y., Friend, D., Ziegler, S. F., Fink, J. S., and Gearing, D. P. Multiple regions within the cytoplasmic domains of the leukemia inhibitory factor receptor and gp130 cooperate in signal transduction in hepatic and neuronal cells. Mol. Cell. Biol. 14, 138-146, 1994.
126. Shabo, Y., Lotem, J., Rubinstein, M., Revel, M., Clark, S. C., Wolf, S. F., Kamen, R., and Sachs, L. The myeloid blood cell differentiation-inducing protein MGI-2A is interleukin 6. Blood 72, 2070-2073, 1988.
127. Miyaura, C., Onozaki, K., Akiyama, Y., Taniyama, T., Hirano, T., Kishimoto, T., and Suda, T. Recombinant human interleukin 6 (B-cell stimulatory factor 2) is a potent inducer of differentiation of mouse myeloid leukemia cells (M1). FEBS Lett 234, 17-21, 1988.(Abstract)
128. Oritani, K., Kaisho, T., Nakajima, K., and Hirano, T. Retinoic acid inhibits interleukin 6-induced macrophage differentiation and apoptosis in a murine hematopoietic cell line, Y6. Blood 80, 2298-2305, 1992.
129. Nakajima, K., Yamanaka, Y., Nakae, K., Kojima, H., Kiuchi, N., Ichiba, M., Kitaoka, T., Fukada, T., Hibi, M., and Hirano, T. A central role for Stat3 in IL-6-induced regulation of growth and differentiation in M1 leukemia cells. EMBO J. 15, 3651-3658, 1996.(Abstract)
130. Minami, M., Inoue, M., wei, S., Takeda, K., Matsumoto, M., Kishimoto, T., and Akira, S. STAT3 activation is a critical step in gp1430-mediated terminal differentiation and growth arrest of a myeloid cell line. Proc. Natl. Acd. Sci. USA. 93, 3963-3966, 1996.
131. Fukada, T., Hibi, M., Yamanaka, Y., Takahashi-Tezuka, M., Fujitani, Y., Yamaguchi, T., Nakajima, K., and Hirano, T. Two signals are necessary for cell proliferation induced by a cytokine receptor gp130: involvement of STAT3 in anti-apoptosis. Immunity. 5, 449-460, 1996.(Abstract)
132. Durbin, J. E., R. Hackenmiller, M. C. Simon, and D. E. Levy. Targeted disruption of the mouse Stat1 gene results in compromised innate immunity to viral disease. Cell 84, 443-450, 1996.
133. Meraz, M. A., J. M. White, K. C. F. Sheehan, E. A. Bach, S. J. Rodig, A. S. Dighe, D. H. Kaplan, J. K. Riley, A. C. Greenlund, D. Campbell, K. Carver-Moore, R. N. DuBois, R. Clark, M. Aguet, and R. D. Schreiber. Targeted disruption of the mouse Stat1 gene in mice reveals unexpected physiologic specificity in the JAK- STAT signaling pathway. Cell 84, 431-442, 1996.
134. Takeda, K., Tanaka, T., Shi, W., Matsumoto, M., Minami, M., Kashiwamura, S., Nakanishi, K., Yoshida, N., kishimoto, T., and Akira, S. Essential role of Stat6 in IL-4 signalling. Nature 380, 627-630, 1996.
135. Shimoda, K., Deursen, J. V., Sangster, M. Y., Sarawar, S. R., Carson, R. T., Tripp, R. A., Chu, C., Quelle, F. W., Nosaka, T. D., Vignali, A. A., Doherty, P. C., Grosveld, G., Paul, W. E., and Ihle, J. N. Lack of IL-4-induced Th2 response and IgE class switching in mice with disrupted Stat6 gene. Nature 380, 630-633, 1996.
136. Kaplan, M. H., Schindler, U., Smiley, S. T., and Grusby, M. J. Stat6 is repuired for mediating responses to IL-4 and for the development of Th2 cells. Immunity 4, 313-319, 1996
137. Thierfelder, W. E., van Deursen, J. M., Yamamoto, K., Tripp, R. A., Sarawar, S. R., Carson, R. T., Sangster, M. Y., Vignali, D. A. A., Doherty, P. C., Grosveld, G. C., and Ihle, J. N. Requirement for Stat4 in interleukin-12-mediated responses of natural killer and T cells. Nature 382, 171-174, 1996.
138. Kaplan, M. H., Sun, Ya-Lin., Hoey, T., and Grusby, M. J. Impaired IL-12 responses and enhanced development of Th2 cells in Stat4-deficient mice. Nature 382, 174-177, 1996.
139. Hou, X. S., Melnick, M. B. and Perrimon, N. Marelle acts downstream of the Drosophila HOP/JAK kinase and encodes a protein similar to the mammalian STATs. Cell 84, 411-419, 1996.
140. Yan, R., Small, S., Desplan, C., Dearolf, C. R., and Darnell, J. E. Jr. Identification of a Stat gene that functions in Drosophila development. Cell 84, 421-430, 1996.
141. Greenlund, A. C., Farrar, M. A., Viviano, B. L., and Schreiber, R. D. Ligand-induced IFN gamma receptor tyrosine phosphorylation couples the receptor to its signal transduction system (p91). EMBO J. 13, 1591-1600, 1994.
142. Hou, J., Schindler, U., Henzel, W. J., Ho, T. C., Brasseur, M., and McKnight, S. L. An interleukin-4-induced transcription factor: IL-4 Stat. Science 265, 1701-1706, 1994.
143. Fujii, H., Nakagawa, Y., Schindler, U., Kawahara, A., Mori, H. Gouilleux, F., Groner, B., Ihle, J. N., Minami, Y., Miyazaki, T., and Taniguchi, T.ハ Activation of Stat5 by interleukin 2 requires a carboxyl-terminal region of the interleukin 2 receptor beta chain but is not essential for the proliferative signal transmission. Proc. Natl.. Acad.. Sci.. 92, 5482-5486, 1995.
144. Friedmann, M. C., Migone, T. S., Russel, S. M., and Leonard, W. J. Different interleukin 2 receptor b-chain tyrosines couple to at least two signaling pathways and synergistically mediate interleukin 2-induced proliferation. Proc. Natl.. Acad.. Sci.. 93, 2077-2082, 1996.
145. Gobert, S., Chretien, S., Gouilleux, F., Muller, O., Pallard, C., Fourt, I. D., Groner, B., Lacombe, C., Gisselbrecht, S., and Mayeux, P. Identification of tyrosine residues within the intracellular domain of the erythropoietin receptor crucial for STAT5 activation. EMBO J. 15, 2434-2441, 1996.
146. Frederic, W. Q., Wang, D., Nosaka, T., Thierfelder, W. E., Stravopodis, D., Weinstein, Y., and Ihle, J. N. Erythropoietin induces activation of Stat5 through association with specific tyrosines on the receptor that are not required for a mitogenic response. Mol. Cell. Biol. 16, 1622-1631, 1996.
147. Mui, A. L., Wakao, H., O'Farrell, A. M., Harada, N., and Miyajima, A. Interleukin-3, granulocyte-macrophage colony stimulating factor and interleukin-5 transduce signals through two STAT5 homologs. EMBO J. 14, 1166-1175, 1995.
148. Yoshikawa, A., Murakami, H., and Nagata, S. Distinct signal transduction through the tyrosine-containing domains of the granulocyte colony-stimulating factor receptor. EMBO J. 14, 5288-5296, 1995.
149. Miyazaki, T., Kawahara, A., Fuji, H., Nakagawa, Y., Minami, Y., Liu, Z. J., Oishi, I., Silvennoinen, O., Witthuhn, B. A., Ihle, J. N., and Taniguchi, T. Functional activation of Jak1 and Jak3 by selective association with IL-2 receptor subunits. Science 286, 1045-1047, 1994.
150. Muller, M., Briscoe, J., Laxton, C., Guschin, D., Ziemiecki, A., Silennoinen, O., Harpur, A. G., Barbieri, G., Witthuhn, B. A., Schindler, C., Pellegrini. S., Wilks, A. F., Ihle, J. N., Stark, G. R., and Kerr, I. M. The protein tyrosine kinase Jak1 complements defects in interferon-a/b and -g signal transduciton. Nature 366, 129-135, 1993.
151. Guschin, D., Rogers, N., Briscoe, J., Witthuhn, B. A., Wathing, D., Horn, F., Pellegrini, S., Yasukawa, K., Heinrich, P., Stark, G. R., Ihle, J. N., and Kerr, I. M. A major role for the protein kinase JAK1 in the JAK/STAT signal transduction pathway in response to interleukin-6. EMBO J. 14, 1421-1429, 1995.
152. Kumar, G., Gupta, S., Wang, S., and Nel, A. E. Involvement of Janus kinases, p52shc, Raf-1, and MEK1 in the IL-6-induced mitogen-activated protein kinase cascade of a growth responsive B cell line. J. Immunol. 153, 4436-4447, 1994.
153. Ernst, M., Gearing, D. P., and Dunn, A. R. Functional and biochemical association of Hck with the LIF/IL-6 receptor signal transducing subunit gp130 in embryonic stem cells. EMBO J. 13, 1574-1584, 1994.
154. Matsuda, T., Takahashi-Tezuka, M., Fukada, T., Okuyama, Y., Funitani, Y., Tshukada, S., Mano, H., Hirai, H., Witte, O. N., and Hirano, T. Association and activation of Btk and Tec tyrosine kinases by gp130, a signal transducer of the interleukin-6 family of cytokines. Blood 85, 627-633, 1995.
155. Matsuda, T., Fukada, T., Takahashi-Tezuka, M., Okuyama, Y., Fujitani, Y., Hanazono, Y., Hirai, H., and Hirano, T. Activation of Fes tyrosine kinase by gp130, an interleukin-6 family cytokine signal transducer, and their association. J. Biol. Chem. 270, 11037-11039, 1995.
156. Takahashi-Tezuka, M., Hibi, M., Fujitani, Y., Fukada, T., Yamaguchi, T.,and Hirano, T. Tec-type tyrosine kinase links the cytokine receptors to PI-3 kinase probably through JAK. Oncogene. 14, 2273-2282, 1997.
157. Hirano, T., Taga, T., Yasukawa, K., Nakajima, K., Nakano, N., Takatsuki, F., Shimizu, M., Murashima, A., Tsunasawa, S., Sakiyama, F., and Kishimoto, T. Human B cell differentiation factor defined by an anti-peptide antibody and its possible role in autoantibody production. Proc. Natl. Acad.. Sci.. USA 84, 228-231, 1987.(Abstract)
158. Hirano, T., Teranishi, T., Toba, T., Sakaguchi, N., Fukukawa, T., and Tsuyuguchi, I. Human helper T cell factor(s) (ThF). I. Partial purification and characterization. J. Immunol. 126, 517-522, 1981.
159. Shoenfeld, Y., and Isenberg, D. A. Mycobacteria and autoimmunity. Immunol. Today 9, 178-182, 1988.
160. Sela, O., E. l.-Roeiy, A., Pick, A. I., and Shoenfeld, Y. Serum immunoglobulin levels in patients with active pulmonary tuberculosis and patients with Klebsiella infection. Immunol. Lett. 15, 117-120, 1987.
161. Isobe, T., and Osserman, E. F. Pathologic conditions associated with plasma cell dyscrasias: A study of 806 cases. Annals. New. York. Acad. Sci.190, 507-517, 1971.
162. Isomaki, H. A., Hakulinen, T., and Joutsenlahti, U. Excess risk of lymphomas, leukemia and myeloma in patients with rheumatoid arthritis. J. Chron. Dis. 31, 691-696, 1978.
163. Rook, G. A. W., and Stanford, J. L. Slow bacterial infections or autoimmunity ? Immunol. Today 13,ハ160-164, 1992.
164. Potter, M., and Boyce, C. Induction of plasma cell neoplasms in strain Balb/c mice with mineral oil and mineral oil adjuvants. Nature 193, 086-1087, 1962.
165. Potter, M., and Wax, J. S. Genetics of susceptibility to pristane-induced plasmacytomas in BALB/cAn: reduced susceptibility in BALB/cJ with a brief description of pristane-induced arthritis. J. Immunol. 127, 1591-1595, 1981.
166. Satoh, M., and Reeves, W. H. Induction of lupus-associated autoantibodies in BALB/c mice by intraperitoneal injection of pristane. J. Exp. Med. 180, 2341-6, 1994.
167. Hirano, T., Matsuda, T., Turner, M., Miyasaka, N., Buchan, G., Tang, B., Sato, K., Shimizu, M., Maini, R., Feldman, M., and Kishimoto, T. Excessive production of interleukin 6/B cell stimulatory factor-2 in rheumatoid arthritis. Eur. J. Immunol. 18, 1797-1801, 1988.(Abstract)
168. Houssiau, F. A., Devogel, J. P., Van Damme, J., De Deuxchaisnes, C. N., and Van Snick, J. Interleukin-6 in synovial fluid and serum of patients with rheumatoid arthritis and other inflammatory arthritides. Arthritis. Rheum.. 31, 784-788, 1988.
169. Bhardwaj, N., Santhanam, U., Lau, L. L., Tatter, S. B., Ghrayeb, J., Rivelis, M., Steinman, R. M., Sehgal, P. B., and May, L. T. ハIL-6/IFN-b2 in synovial effusions of patient with rheumatoid arthritis and other arthritides. J. Immunol. 143, 2153-2159, 1988.
170. Hermann, E., Fleischer, B., Mayer, W. J., Poralla, T., and Meyer Zum Beschernfelde, K. H.Correlation of synovial fluid interleukin 6 (IL-6) antivities with IgG concentrations in patients with inflammatory joint disease and osteoarthritis. Clin. Exp. Rheumatol.7, 411-414, 1989.
171. Guerne, P. A., Zuraw, B. L., Vaughan, J. H., Carson, D. A., and Lotz, M. Synovium as a source of interleukin 6 in vitro: contribution to local and systemic manifestations of arthritis. J Clin Invest 83, 585-592, 1989.
172. Shinmei, M., Masuda, K., Kikuch, T., and Shimomura, Y. The role of cytokines in chondrocyte mediated cartilage degradation. J. Rheumatol. 16, 32-34, 1989.
173. Takai, Y., Seki, N., Senoh, H., Yokota, T., Lee, F., Hamaoka, T., and Fujiwara, H. Enhanced production of interleukin-6 in mice with type II collagen-induced arthritis. Arthritis. Rheum.. 32, 594-600, 1989.
174. Tang, B., Matsuda, T., Akira, S., Nagata, N., Ikehara, S., Hiranio, T., and Kishimoto, T. Age-associated increase in interleukin-6 in MRL/lpr mice. International Immunol. 3, 273-278, 1991.
175. Taga, T., Kawanishi, K., Hardy, R. R, Hirano, T., and Kishimoto, T. Receptors for B cell stimulatory factor 2 ( BSF-2 ): Quantitation, Specificity, Distribution and Regulation of the Expression. J. Exp. Med. 166, 967-981, 1987.
176. Tanaka, Y., Saito, K., Shirakawa, F., Ota ,T., Suzuki, Sumiya, H. E., and Yamashita, Y. Production of B cell-stimulating factors by B cells in patients with systemic lupus erythematosus. J. Immunol. 141, 3043-3049, 1988.
177. Klashman, D. J., Martin, R. A., Martinez-Maza, O., and Stevens, R. H. In vitro regulation of B cell differentiation by interleukin-6 and soluble CD23 in systemic lupus erythematosus B cell subpopulations and antigen-induced normal B cells. Arthritis and Rheumatism 34, 276-286, 1991.
178. Nagafuchi, H., Suzuki, N., Mizuchima, Y., and Sakane, T. constitutive expression of iL-6 receptors and their role in the excessive B cell function in patients with systemic lupus erythematosus. J. Immunol. 151, 6525-6534, 1993.
179. Kobayashi, I., Matsuda, T., Saito, T., Yasukawav, K., Kikutani, H., Hirano, T., Taga, T., and Kishimoto, T. Abnormal distribution of IL-6 receptor in aged MRL/lpr mice: elevated expression on B cells and absence on CD4+ cells. Inter. Immunol. 4, 1407-1412, 1992.
180. Kanno, K., Okada, T., Abe, M., Hirose, S., Shirai. T. Differential sensitivity to interleukins of CD5+ and CD5- anti-DNA antibody-producing B cells in murine lupus. Autoimmunity 14, 205-214, 1993.
181. Ryffel, B., Car, B. D., Gunn, H., Roman, D., Hiestand, P., and Mihatsch, M. J. Interleukin-6 exacerbates glomerulonephritis in (NZB x NZW)F1 mice. Am. J. Pathol.144, 927-37, 1994.
182. Bendtzen, K., Buschard, K., Diamant, M., Horn, T., and Svenson, M. Possible role of IL-1, TNF-alpha, and IL-6 in insulin-dependent diabetes mellitus and autoimmune thyroid disease. Lymphokine Res. 8, 335-340, 1989.
183. Campbell, I. L., Cutri, A., Wilson, A., and Harrison, L. C. Evidence for IL-6 production by and effects on the pancreatic beta-cell. J. Immunol. 143, 1188-1191, 1989.
184. Campbell, I. L., and Harrison, C. A new view of the beta cell as an antigen-presenting cell and imunogenic target. J. Autoimmunity 3, 53-62, 1990.
185. Rook, G. A.W., Thompson, S., Buckley, M., Elson, C., Brealey, R., Lambert, C., White, T., and Rademacher, T. The role of oil and agalactosyl IgG in the induction of arthritis in rodent models. Eur. J. Immunol. 21, 1027-1032, 1991.
186. Nakao, H., Nishikawa, A., Nishiura, T., Kanayama, Y., Tarui, S., and Taniguchi, N. Hypogalactosylation of immunoglobulin G sugar chains and elevated serum interleukin 6. Clin. Chim. Acta. 197, 221-228, 1991.
187. Campbell, I. L., Hobbs, M. V.,and Dockter, J. Oldstone-MB; Allison-JIslet inflammation and hyperplasia induced by the pancreatic islet-specific overexpression of interleukin-6 in transgenic mice. Am. J. Pathol. 145, 157-66, 1994.
188. Campbell, I. L., Kay, T. W., Oxbrow, L., and Harrison, L. C. Essential role for interferon-g and interleukin-6 in autoimmumne insulin-dependent diabetes in NOD/wehi mice. J. Clin. Invest. 87, 739-742, 1991.
189. Finck, B., A, K., Chan, B., and Wofsy, D. Interleukin 6 promotes murine lupus in NZB/NZW F1 mice. J. Clin. Invest.. 94, 585-91, 1994.
190. Potter, M. Genetics of susceptibility to plasmacytoma development in BALB/c mice. Cancer Surveys 3, 247-264, 1984.
191. Jacob, C. O., and McDevitt, H. O., Genetic predisposition to autoimmune diseases: the contribution of the major histocompatibility complex. In: Molecular Autoimmunity 1991, edited by Talal, N. London: Academic Press, pp. 7-29.
192. Jacob, C. O., and McDevitt, H. O. Interferon gamma and tumor necrosis factor in autoimmune disease models: implications for immunoregulation and genetic susceptibility. In: Molecular Autoimmunity 1991, edited by Talal, N. London: Academic Press, pp. 107-126.
193. Watanabe-Fukunaga, R., Brannan, C. I., Copeland, N. G., Jenkins, N. A., and Nagata, S. Lymphoproliferation disorder in mice explained by defects in Fas antigen that mediates apoptosis. Nature 256, 314-317, 1992.
194. Takahashi, T., Tanaka, M., Brannan, C. I., Jenkink, N. A., Copeland, N. G., Suda, T., and Nagata, S. Generalized lymphoproliferative disease in mice, caused by a point mutation in the Fas ligand, Cell 76, 969-976, 1994.
195. Hirano, T. Interleukin-6 and its relation to inflammation and disease. Clinical Immunol. and Immunopathol 62, S60-S65, 1992.(PubMed)
196. Hirano, T. Interleukin 6 and autoimmunity and plasma cell neoplasias. Res. Immunol. 143, 759-763, 1992.
197. Hirano, T. Interleukin 6 transgenic mouse: A model of chronic inflammatory proliferative disease. In: Overexpression and knockout of cytokines in transgenic mice. 1994, edited by Jacob, C. O., London: Academic Press, pp. 73-84.
198. Kaisho, T., Oritani, K., Ishikawa, J., Tanabe, M., Muraoka, O., Ochi, T., and Hirano, T. Human bone marrow stromal cell lines from myeloma and rheumatoid arthritis which can support murine preB cell growth. J. Immunol 149, 4088-4095, 1992.
199. Kaisho, T., Ishikawa, J., Oritani, K., Inazawa. J., Tomizawa, H., Muraoka, O., Ochi, T., and Hirano, T. BST-1, a surface molecule of bone marrow stromal cell lines that facilitates pre-B cell growth. Proc. Natl. Acad. Sci .USA 91, 5325-5329, 1994.(Abstract)
200. Garry, R. F., Fermin, C. D., Hart, D. J., Alexander, S. S., Donehower, L. A., Lou-Zhang, H. Detection of a human intracisternal A-type retroviral particle antigenically related to HIV. Science 250, 1127-1129, 1990.
201. Trabandt, A., Gay, R. E., Stransky, G. , Aicher. W. K., and Gay, S. Cellular and molecular basis of rheumatoid joint destruction. In: Molecular Autoimmunity 1991, edited by Talal, N., London: Academic Press, pp 233-245.
202. Kitajima, I., Yamamoto, K., Sato, K., Nakajima, Y., Nakajima, T., Maruyama, I., Osame, M., and Nishioka, K. Detection of human T cell lymphotropic virus type I proviral DNA and its gene expression in synovial cells in chronic inglammatory arthropathy. J. Clinical Investigation 88, 1315-1322, 1991.
203. Iwakura, Y., Tosu, J., Yoshida, E., Takiguchi, M., Sato, K., Kitajima, I., Nishioka, K., Yamamoto, K., Takeda, T., Hatanada, M., Yamamoto, H., and Sekiguchi, T. Induction of inflammatory arthropathy resembling rheumatoid arthritis in mice transgenic for HTLV-I. Science 253, 1026-1028, 1991.
204. Inoue , J. I., Seiki, M., Taniguchi, T., Tsuru, S., and Yoshida, M. Induction of interleukin 2 receptor gene expression by p40x encoded by human T-cell leukemia virus type 1. EMBO J. 5, 2883-2888, 1986.
205. Maruyama, M., Shibuya, H., Harada, H., Hatakeyama, M., Seiki, M., Fujita, T., Inoue, J., Yoshida, M., and Taniguchi, T. Evidence for aberrant activation of the interleukin-2 autocrine loop by HTLV-I ecnoded p40x and T3/Ti complex triggering. Cell 48, 343-350, 1987.
206. Cross, S. L., Feinberg, M. B., Wolf, J. B., Holbrook, N. J., Wong-Staal, F., and Leonard, W. J. Regulation of the human interleukin-2 receptor chain promoter: activation of a nonfunctional promoter by transactivator gene of HTLV-I. Cell 49, 47-56, 1987.
207. Arai, N., Nomura, D., Villaret, D., Malefijt, R. D., Seiki, M., Yoshida, M., Minoshima, S., Fukayama, R., Maekawa, M., Kudoh, J., Shimizu, N., Yokota, K., Abe. E., Yokota, T., Takebe, Y., and Arai, K. Complete nucleotide sequence of the chromosomal gene for human IL-4 and its expression. J. Immunol. 142, 274-282, 1989.
208. Nimer, S. D., Gasson, J. C., Hu, K., Smalberg, I., Williams, J. L., Chen, I. S., and Rosenblatt, J. D. Activation of the GM-CSF promoter by HTLV-1 and -II tax proteins. Oncogene 4, 671-676, 1989.
209. Kim, S. J., Kehrl, J. H., Burton, J., Tendler, C. L., Jeang, K. T., Danielpour, D., Thevenin, C., Kim, K. Y., Sporn, M. B., and Robert, A. B. Transactivation of the transforming growth factorb1 (TGF-b1) gene by human T lymphotropic virus type 1 tax: A potential mechanism for the increased production of TGF-b1 in adult T cell leukemia. J. Exp. Med. 172, 121-129, 1990.
210. Martinez-Maza, O. IL-6 and AIDS. Res. Immunol.143, 764-769, 1992.
211. Muraoka, O., Kaisho, T., Tanabe, M., and Hirano, T. Transcriptional activation of the interleukin 6 gene by HTLV-1 p40tax through an NF-kB-like binding site. Immunol. Letter 37, 159-165, 1993.
212. Armstrong, M. Y. K., Ebenstein, P., Konigsberg, W. H., and Richards, F. F. Endogenous RNA tumor viruses are activated during chemical induction of murine plasmacytomas. Proc. Natl. Acad.. Sci.. USA 75, 4549-4552, 1978.
213. Feghali, C. A., Bost, K. L., Boulware, D. W., and Levy, L. S. Control of IL-6 expression and response in fibroblasts from patients with systemic sclerosis. Autoimmunity 17(4), 309-18, 1994.
|
|
|