 |
|
||
 |
|


 |
| HOME 亜 尋媶偺曽岦惈偲彨棃揥朷 |
崱屻偺尋媶偺曽岦惈偲彨棃揥朷惗壔妛(Biochemistry)偼惗暔傪壔妛揑側帇揰偱棟夝偟傛偆偲偡傞妛栤偺懱宯偱偡丅惗懱撪偺抈敀幙丄摐幙丄帀幙傪扨棧偟丄偦傟傜偺暔幙曄姺偲僄僱儖僊乕戙幱偺奣梫偑1950擭戙拞崰傑偱偵柧傜偐偵側傝傑偟偨丅1953擭偵僲乕儀儖堛妛惗棟妛徿偑H.A. Krebs攷巑偲F.A. Lipman攷巑偺乽僩儕僇儖儃儞巁僒僀僋儖偺敪尒偲戙幱偵偍偗傞崅僄僱儖僊乕儕儞巁寢崌偺堄媊偺敪尒乿偵懳偟偰梌偊傜傟偨偺偼丄偦偺堦偮偺惉壥偱偟傚偆丅嫽枴怺偄偙偲偵丄摨擭偵J.D. Watson攷巑偲F.H. Click攷巑偵傛傝DNA擇廳傜偣傫儌僨儖偑採彞偝傟丄偦偺屻暘巕惗暔妛偺媄弍揑恑曕偲憡傑偭偰丄1970擭戙屻敿偵偼妀巁傕帺桼偵埖偊傞帪戙偑摓棃偟傑偟偨丅惗懱傪峔惉偡傞庡梫側4庬椶偺桳婡壔崌暔乮抈敀幙丄摐幙丄帀幙丄妀巁乯偺峔憿偲婡擻傪棟夝偡傞偙偲偵傛傝丄惗柦傪棟夝偟傛偆偲偡傞帋傒偑側偝傟丄惗壔妛丄惗棟妛丄嵶朎惗暔妛丄夝朥妛摍偲丄庡偲偟偰尋媶庤朄偵傛傝暘椶偝傟偰偄偨妛栤椞堟偺暻偼嬌傔偰掅偔側傝丄惗柦壢妛偲偄偆奣擮偵廤栺偝傟偰偄偒傑偟偨丅傑偝偵丄20悽婭偺嵟屻偺20擭娫偵旘桇揑偵恑曕偟偨惗柦壢妛偼丄僸僩堚揱巕抧恾偺嶌惢乹僎僲儉僾儘僕僃僋僩乺偵戙昞偝傟傞傛偆偵丄娨尦揑傾僾儘乕僠偑庡棳偱偁傝丄嵶朎傪偦偺峔惉梫慺偵暘夝偟偰棟夝偡傞偙偲偱偟偨丅巹払偼丄1980擭戙屻敿偐傜1990擭戙慜敿偵偐偗偰惗懱慻怐偐傜悢乆偺掅暘巕検G抈敀幙偲偦偺妶惈惂屼抈敀幙傪惛惢偟丄偦偺墫婎攝楍傪寛掕偟丄東栿屻廋忺傗婡擻傪柧傜偐偵偟偰偒傑偟偨丅偝傜偵丄1990擭戙屻敿偐傜2000擭戙慜敿偵偐偗偰偼丄Wnt娭楢抈敀幙傪摨掕偟丄偦偺婡擻夝愅傪峴偭偰偒傑偟偨丅偦傟偑嵶朎偺僔僌僫儖揱払惂屼傗憹怋丄塣摦丄彫朎桝憲偺暘巕婡峔偺棟夝偵峷專偟偨偙偲偼帠幚偱偡丅 偟偐偟丄嵶朎傪偦偺峔惉梫慺偐傜嵞峔惉偡傞応崌丄慡懱偼昁偢偟傕晹暘偺憤榓偱側偔丄暘巕娫偺憡屳嶌梡偺寢壥丄怴偨側惈幙傪妉摼偟傑偡丅偝傜偵丄屄懱傪棟夝偡傞偨傔偵偼丄嵶朎偲嵶朎偺楢実丄憻婍偲憻婍偺楢実傪峫偊側偔偰偼側傝傑偣傫乮恾侾乯丅屄乆偺暘巕傪夝愅偟偰偦偺婡擻傪柧傜偐偵偟偰偄偔尋媶庤朄偺廳梫惈偼曄傢傝傑偣傫偑丄巹払偼偙傟傑偱偵攟偭偰偒偨惛鉱側尋媶庤朄傪梡偄偰怴偨側尋媶暘栰偵僠儍儗儞僕偟偰偄偙偆偲峫偊偰偄傑偡丅惗壔妛揑側庤朄偲峫偊曽傪朰傟傞偙偲側偔丄傛傝峀偄帇栰偱暔帠傪峫偊丄忢偵幘姵偲偺娭楢傪擮摢偵抲偒丄尋媶傪揥奐偟傑偡丅More Biochemistry丄More Biology丄More Diseases偑僉儍僢僠僼儗乕僘偱偡丅彮偟嬶懱揑偵弎傋傑偟傚偆丅 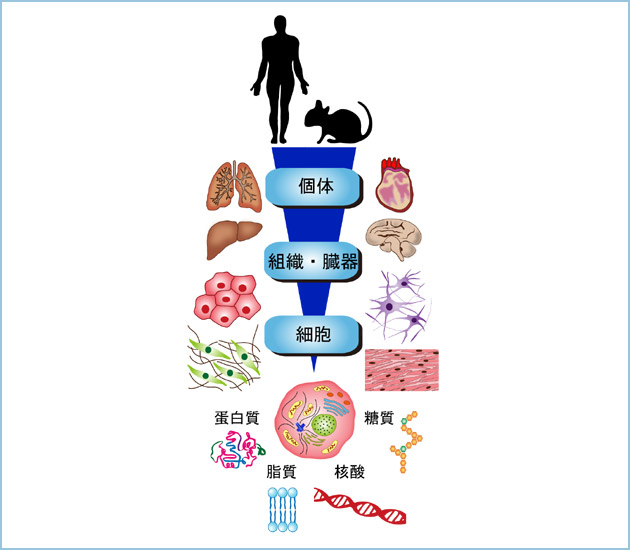 恾侾丏屄懱傪峔惉偡傞梫慺偲偦偺楢実
屄懱偼慻怐丒憻婍偐傜丄慻怐丒憻婍偼嵶朎偐傜丄傑偨嵶朎偼4庬椶偺庡梫側桳婡壔崌暔偐傜峔惉偝傟偰偄傞丅屄乆偺峔惉梫慺偼屳偄偵楢実偟丄憡屳嶌梡偡傞偙偲偱怴偨側惈幙傪妉摼偟偰偄偔丅
More Biochemistry
忋弎偟傑偟偨傛偆偵丄惗柦尰徾偵晄壜寚側抈敀幙丄堚揱巕乮妀巁乯丄帀幙丄摐幙摍偺峔憿偲婡擻偺娭學丄偦傟傜偺戙幱傗挷愡婡峔偑師乆偵夝柧偝傟偰偒傑偟偨丅摿偵抈敀幙偵娭偡傞尋媶偼丄堚揱巕岺妛偺敪揥偵敽偄栚揑偺抈敀幙敪尰偑壜擻偲側偭偨寢壥丄旘桇揑偵恑揥偟傑偟偨丅20悽婭傑偱偺惗柦壢妛偺尋媶懳徾偺庡棳偼抈敀幙偐傜堚揱巕傊偲曄傢偭偰偒傑偟偨偑丄2003擭偵僸僩僎僲儉夝愅偑廔傢傝丄嵞傃抈敀幙傊偲堏傝偮偮偁傝傑偡丅僎僲儉偵僐乕僪偝傟偨堚揱巕偼mRNA偵揮幨偝傟偨屻丄儕儃僜乕儉偵偍偄偰東栿偝傟抈敀幙偑惗崌惉偝傟傑偡偑丄懡偔偺抈敀幙偼億儕儁僾僠僪偲偟偰崌惉偝傟偨屻偵丄儕儞巁壔傗摐幙丄帀幙摍偺東栿屻廋忺偲懠偺抈敀幙偲暋崌懱宍惉偡傞偙偲偵傛傝丄惗棟婡擻偑惂屼偝傟傑偡丅偙偺堚揱忣曬偩偗偱偼暘偐傜側偄抈敀幙偺惗壔妛揑惈忬傪夝愅偟丄偳偺傛偆側暘巕婡峔偱妶惈偑惂屼偝傟偰惗棟婡擻傪敪婗偟偰偄傞偺偐傪夝柧偡傞偙偲偑偙傟偐傜偺惗柦壢妛尋媶偵偍偄偰廳梫側壽戣偱偡丅偟偨偑偄傑偟偰丄幙検暘愅朄偺奿抜偺恑曕偵敽偭偰丄惗壔妛揑夝愅偺廳梫惈偼崱屻傑偡傑偡憹戝偡傞傕偺偲峫偊傜傟傑偡丅 懡嵶朎惗暔偺弶婜敪惗傗峆忢惈偺堐帩偼丄嵶朎偺憹怋傗暘壔丄塣摦丄愙拝丄嬌惈摍偺嵶朎墳摎偑憡屳偵楢娭偡傞偙偲偵傛傝惂屼偝傟偰偄傑偡丅偙傟傜偺懡庬懡條側嵶朎墳摎偼丄BMP傗TGF丄EGF丄FGF丄Wnt摍偺嵶朎奜暘斿宆憹怋暘壔場巕偵傛傝惂屼偝傟偰偄傑偡丅儂儖儌儞傗憹怋暘壔場巕偵傛傝惂屼偝傟傞嵶朎婡擻傪柧傜偐偵偡傞尋媶偺墹摴偼丄傑偢偙傟傜偺抈敀幙傗儁僾僠僪傪惛惢偡傞偙偲偱偡丅椺偊偽丄1977擭偵乽擼偺儁僾僠僪儂儖儌儞惗嶻偵娭偡傞敪尒乿偱僲乕儀儖堛妛惗棟妛徿傪庴徿偟偨R. Gilman攷巑偲A.V. Schally攷巑偼悢廫乣昐枩摢暘偺撠傗梤偺帇彴壓晹偐傜TRF乹峛忬態巋寖儂儖儌儞暘斿場巕乺傪惛惢偟傑偟偨丅巹払偼丄尰嵼Wnt僔僌僫儖傪尋媶偺拞怱偵悩偊偰偄傑偡偑丄僸僩偵偍偄偰19庬椶懚嵼偡傞Wnt偼僔儑僂僕儑僂僶僄傗傾僼儕僇僣儊僈僄儖丄儅僂僗傪梡偄偨堚揱妛揑夝愅偵傛傝丄弶婜敪惗偺懱幉傗懱愡丄婍姱宍惉偵偍偗傞偦偺廳梫惈偑柧傜偐偵偝傟偰偒傑偟偨丅偟偐偟丄Wnt尋媶偵偍偗傞忈暻偺堦偮偵惗棟妶惈傪桳偡傞抈敀幙偲偟偰Wnt傪扨棧丄惛惢偡傞偙偲偑崲擄偱偁偭偨偙偲偑嫇偘傜傟傑偡丅偦傟偼丄Wnt偑摐幙傗帀幙偵傛傞廋忺傪庴偗偰偍傝丄晄梟惈偱幐妶偟堈偔丄嵶朎奜儅僩儕僢僋僗偵寢崌偡傞孹岦偑嫮偄偐傜偱偡乮恾俀乯丅偦偺偨傔偵丄Wnt僔僌僫儖傪妶惈壔偡傞偵偼嵶朎偵Wnt偺cDNA傪夁忚敪尰偝偣傞偐丄Wnt抈敀幙傪娷傫偩攟梴忋惔傪嶌梡偝偣傞偟偐偱偒傑偣傫偱偟偨丅偙傟傜偺幚尡庤朄偱偼丄Wnt杮棃偺妶惈傪尒弌偡偙偲偼偱偒傑偣傫丅2003擭偵Wnt3a偺惛惢朄偑曬崘偝傟丄巹払傕偦偺曽朄傪夵椙偡傞偙偲偵傛傝丄Wnt3a偵壛偊偰Wnt5a丄Wnt5b偺惛惢偵惉岟偟偰偄傑偡丅Wnt偺懡條側惗棟妶惈傪峫偊傟偽丄崱屻偼忋婰埲奜偺Wnt抈敀幙偺惛惢朄傕妋棫偟丄偦傟傜偺惛惢Wnt抈敀幙傪梡偄偰丄乮1乯摐嵔傗帀幙偺東栿屻廋忺傪夘偡傞Wnt偺嵶朎奜暘斿偺惂屼丄乮2乯庴梕懱偲偺摿堎揑寢崌偵傛傞暋悢懚嵼偡傞Wnt僔僌僫儖宱楬偺慖戰揑妶惈壔丄乮3乯嵶朎撪偵偍偗傞抈敀幙娫憡屳嶌梡傗儕儞巁壔偵傛傞Wnt僔僌僫儖偺妶惈壔婡峔傪柧傜偐偵偡傞偙偲偑Wnt僔僌僫儖偵傛傞嵶朎墳摎偺慡杄傪柧傜偐偵偡傞忋偱昁梫偲峫偊偰偄傑偡丅傑偨丄暔幙傪廳梫帇偡傞惗壔妛偺揱摑揑側峫偊曽傪朰傟側偄傛偆偵偟偨偄偲巚偭偰偄傑偡丅 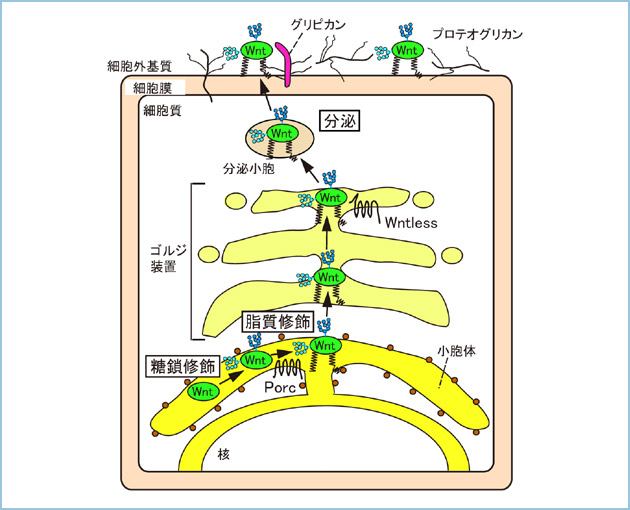 恾俀丏Wnt偺東栿屻廋忺偲嵶朎奜暘斿偺惂屼
Wnt偼彫朎懱偲僑儖僕懱偵偍偄偰摐嵔廋忺傗Porcupine乮Porc乯偵傛傞帀幙廋忺傪庴偗傞偲妶惈宆偲側傞丅偦偺屻丄Wntless偵傛傝嵶朎奜偵暘斿偝傟偨Wnt偼僾儘僥僆僌儕僇儞摍偺嵶朎奜婎幙偺峔惉暘巕偲寢崌偟丄嵶朎奜桝憲傗僔僌僫儖揱払偑惂屼偝傟傞丅
More Biology
惗暔傪峔惉偟丄懡嵤側惗柦尰徾傪惗傒弌偟偰偄傞嵟彫偺扨埵偼嵶朎偱偡丅偦偺偨傔傑偢柧傜偐偵偟側偗傟偽側傜側偄偺偼屄乆偺嵶朎偑偳偺傛偆偵偦偺婡擻傪惂屼偟偰偄傞偺偐偲偄偆栤戣偱偡丅嵶朎偼嵶朎奜偐傜懡嵤側忣曬傪庴偗庢傝丄嵶朎撪偺僔僌僫儖揱払傪妶惈壔偡傞偙偲偱堚揱巕敪尰傗憹怋丄暘壔丄塣摦丄嬌惈壔摍偺嵶朎墳摎傪偟傑偡(恾俁)丅巹払偼偙傟傑偱憹怋暘壔場巕傪拞怱偲偟偨嵶朎奜忣曬偑嵶朎撪僔僌僫儖揱払宱楬傪妶惈壔偟丄嵶朎墳摎傪堷偒婲偙偡巇慻傒傪夝愅偟偰偒傑偟偨丅摿偵丄偙傟傑偱惛惢偑擄偟偄偲偝傟偰偄偨Wnt摍偺憹怋暘壔場巕傪惛惢偟丄嵶朎偵嶌梡偝偣傞偙偲偱偦偺惗棟揑側妶惈傗僔僌僫儖揱払婡峔傪暘巕儗儀儖偱徻嵶偵柧傜偐偵偟偰偒傑偟偨丅 偟偐偟丄巹払偑崱屻丄傛傝惗棟揑側嵶朎婡擻傪柧傜偐偵偟偰偄偔忋偱朰傟偰偼側傜側偄偺偼懡偔偺嵶朎偑屄懱偵偍偄偰偼廤抍偲偟偰慻怐傗憻婍傪宍偯偔傝丄屌桳偺婡擻傪敪婗偟偰偄傞偲偄偆偙偲偱偡丅嵶朎偼廤抍偲偟偰慻怐壔偡傞偙偲偱偼偠傔偰惗棟揑側宍懺傗嬌惈丄暘壔忬懺摍傪妉摼偡傞偙偲偼傛偔抦傜傟偰偄傑偡丅偦偙偱巹払偼尰嵼丄塼惈場巕傗愙拝偲偄偭偨嵶朎奜僔僌僫儖偑丄慻怐壔偟偨嵶朎廤抍偵懳偟偰偳偺傛偆側墳摎傪堷偒婲偙偡偺偐偵偮偄偰嫽枴傪傕偭偰尋媶傪恑傔偰偄傑偡丅嵶朎偑嵟傕妶敪偵慻怐壔偟偰嵶朎奜僔僌僫儖偵墳摎偟丄婡擻傪敪婗偡傞偺偑屄懱敪惗夁掱偱偡丅偦偙偱巹払偺尋媶幒偱偼暋悢偺憹怋暘壔場巕偵偮偄偰僲僢僋傾僂僩儅僂僗傪嶌惢偟丄敪惗夁掱偵偍偗傞惗棟揑側栶妱傪徻嵶偵専摙偟偰偄傑偡丅傑偨丄慻怐傗憻婍摿堎揑偵堚揱巕傪寚懝偝偣傞僐儞僨傿僔儑僫儖僲僢僋傾僂僩儅僂僗偺媄弍傕墳梡偟丄摿掕偺嵶朎庬丄傑偨慻怐偵偍偗傞婡擻夝愅傕愊嬌揑偵峴偭偰偄傑偡丅 歁擕摦暔偺屄懱敪惗偼曣懱撪偱恑峴偡傞偨傔偦偺夁掱傪娤嶡偡傞偙偲偑擄偟偔丄傑偨旕忢偵懡庬椶偺嵶朎偑暋嶨偐偮嫤挷揑偵摥偄偰惂屼偝傟偰偄傞偨傔丄惗壔妛傗嵶朎惗暔妛揑側暘巕儗儀儖偱偺夝愅偑崲擄偱偁傞偙偲偼傛偔抦傜傟偰偄傑偡丅偦偙偱巹払偼婍姱攟梴傗嶰師尦攟梴偲偄偭偨媄弍傪梡偄偨夝愅傪峴偭偰偄傑偡乮恾係乯丅嬶懱揑偵偼丄儅僂僗戀帣偐傜攛傗恡憻丄懥塼態側偳偺憻婍尨婎傪庢傝弌偟偰in vitro偱婍姱攟梴偟丄摿掕偺憹怋暘壔場巕傪嶌梡偝偣傞偙偲偱慻怐丒憻婍偺敪惗夁掱偵偍偗傞嶌梡傪栚偵尒偊傞宍偱専摙偟偰偄傑偡丅傑偨僲僢僋傾僂僩儅僂僗偐傜庢傝弌偟偨憻婍尨婎傪婍姱攟梴偟丄敪惗夁掱傪栰惗宆偲斾妑偡傞偙偲偱嵟廔揑側昞尰宆偵帄傞傑偱偺夁掱傪徻嵶偵夝愅偟偰偄傑偡丅偝傜偵憻婍偐傜娫幙嵶朎傪彍偄偰忋旂嵶朎廤抍偺傒傪庢傝弌偟丄嵶朎奜婎幙僎儖撪偱摿掕偺場巕偺懚嵼壓偱嶰師尦攟梴偡傞曽朄偱傕慻怐敪惗夁掱傪嵞尰偡傞偙偲偑偱偒傑偡丅偙偺媄弍傪墳梡偟偰屄懱慡懱偱偼夝愅偡傞偙偲偑擄偟偄摿掕偺嵶朎廤抍偵懳偡傞憹怋暘壔場巕偺嶌梡傪専摙偟偰偄傑偡丅嵶朎愼怓傗惗壔妛揑庤朄偵傛傝摿掕偺抈敀幙傗堚揱巕偺敪尰傗嬊嵼丄妶惈壔忬懺偺曄壔傪偙傟傜偺攟梴媄弍偲慻傒崌傢偣偰夝愅偟丄惗棟揑側嵶朎偺嬌惈丄塣摦傗暘壔偺惂屼儊僇僯僘儉傪暘巕儗儀儖偱柧傜偐偵偡傞偙偲傪栚巜偟傑偡丅 偙偺傛偆偵丄巹払偼屄乆偺嵶朎儗儀儖偐傜慻怐傗憻婍丄偦偟偰屄懱儗儀儖偵帄傞傑偱摑崌揑偵惗柦尰徾傪棟夝偡傞偲偄偆巔惃傪戝愗偵偟側偑傜擔乆尋媶傪恑傔偰偄傑偡丅僸僩傪偼偠傔偲偡傞懡嵶朎惗暔偑偳偺傛偆偵偟偰帺傜偺屄懱傪宍偯偔傝丄傑偨偦傟傪堐帩偟偰偄傞偐丄偦偺惂屼儊僇僯僘儉傪惗壔妛傗嵶朎惗暔妛揑庤朄傪嬱巊偟偰暘巕儗儀儖偱柧傜偐偵偡傞偙偲偑戝偒側僥乕儅偺堦偮偱偡丅 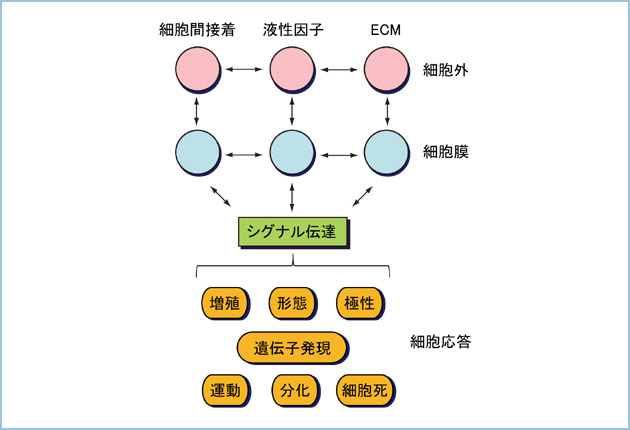 恾俁丏懡嵤側嵶朎奜巋寖偲嵶朎墳摎
嵶朎偼嵶朎娫愙拝丄塼惈場巕傗ECM乮嵶朎奜儅僩儕僢僋僗乯偲偄偭偨懡嵤側嵶朎奜巋寖傪庴偗庢傝丄嵶朎撪僔僌僫儖揱払宱楬傪妶惈壔偝偣傞偙偲偱堚揱巕敪尰傗憹怋丄暘壔丄塣摦摍偺條乆側嵶朎墳摎傪偡傞丅
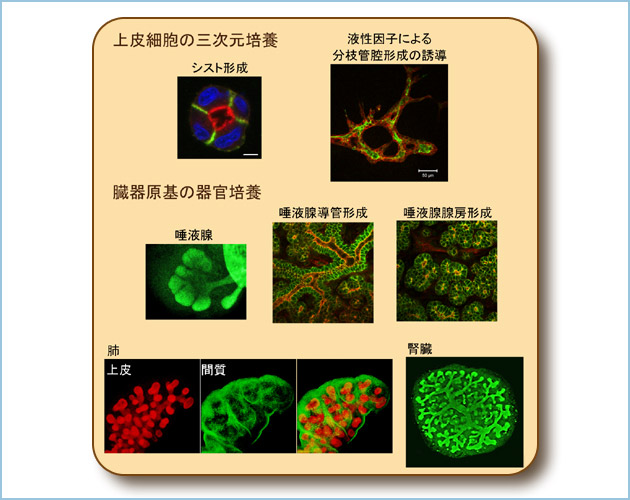 恾係丏嶰師尦攟梴丒婍姱攟梴傪梡偄偨嵶朎慻怐壔偺惂屼儊僇僯僘儉偺夝愅
忋旂嵶朎偺嶰師尦攟梴傪峴偄丄撪峯偲捀掙嬌惈傪桳偟偨僔僗僩宍惉傗丄塼惈場巕偵傛傞暘巬娗峯宍惉偺夝愅傪峴偆丅儅僂僗戀帣偐傜敪惗夁掱偺憻婍尨婎傪庢傝弌偟丄婍姱攟梴偡傞偙偲偱徻嵶側婍姱宍惉夁掱偲偦偺惂屼儊僇僯僘儉傪暘巕儗儀儖偱柧傜偐偵偡傞丅
More Diseases
巹払偼偙傟傑偱丄娻偺敪徢傗揮堏偺巇慻傒傪夝柧偡傞偨傔偺尋媶傪峴偭偰偒傑偟偨丅1970乣90擭戙丄亀娻堚揱巕亁傗亀娻梷惂堚揱巕亁偲屇偽傟傞堚揱巕偑婔偮傕摨掕偝傟丄偦偺寢壥丄乽僂僀儖僗敪娻乿傗乽壔妛敪娻乿丄乽曻幩慄敪娻乿摍偺奣擮偼摑堦偝傟丄娻偼僎僲儉偵偍偗傞僟僀僫儈僢僋側曄壔偵敽偆幘姵偱偁傝丄偦偺婎杮偼桪惃偺婡擻妉摼傪偟偨娻堚揱巕嶻暔偲婡擻偑楎惃憆幐偝傟偨娻梷惂堚揱巕嶻暔傪嶻惗偡傞撍慠曄堎偑暋崌偟偨傕偺偱偁傞偲峫偊傜傟傞傛偆偵側傝傑偟偨丅偙偺偙偲偼崅昿搙偵曄堎傪堷偒婲偡傛偆側梫場偵憳嬾偟偨傝丄傕偟偔偼娻娭楢堚揱巕偺曄堎傪愭揤揑偵桳偟偰偄傞応崌偵偼丄娻偑敪徢偡傞壜擻惈偑崅偔側傞偙偲傪堄枴偟傑偡丅傑偨丄娻堚揱巕偑僐乕僪偡傞抈敀幙偵偼丄憹怋暘壔場巕傗偦偺嵶朎撪僔僌僫儖揱払場巕偑悢懡偔娷傑傟丄憹怋暘壔場巕偺僔僌僫儖揱払婡峔偺攋抅偑娻偲枾愙偵娭楢偡傞偙偲傕柧傜偐偵側偭偰偒傑偟偨丅偟偐偟丄慡偰偺娻偑丄偙偺傛偆側堚揱巕偺曄堎偺拁愊偵傛偭偰偺傒敪徢偡傞傢偗偱偼偁傝傑偣傫丅嵟嬤偱偼丄嵶朎傪庢傝姫偔娐嫬梫場偑丄娻偺敪徢傗恑峴偵娭梌偡傞偙偲偑拲栚偝傟偰偄傑偡丅偲傝傢偗枬惈揑側亀墛徢亁偲偺娭楢偑拲栚偝傟偰偄傑偡丅 墛徢偲偼惗懱偑彎奞傪庴偗偨帪偵婲偙偡斀墳偱丄嵶嬠摍偺奜晹偐傜偺怤擖暔偵懳偡傞柶塽墳摎偑偦偺揟宆偱偡丅偙偺応崌丄T嵶朎傗B嵶朎丄儅僋儘僼傽乕僕摍偺柶塽扴摉嵶朎偑彎奞晹埵偵尰傟偰丄僒僀僩僇僀儞傪暘斿偡傞偙偲偵傛傝丄柶塽扴摉嵶朎偺憹堳傗屳偄偺嵶朎巰偺梷惂丄偝傜偵偼僒僀僩僇僀儞暘斿傪懀恑偝偣傞偙偲偱堦憌杊屼擻傪嫮傔傑偡丅堦曽丄僒僀僩僇僀儞偺嬊強揑側憹壛傪夘偟偨偙偺傛偆側尰徾偑丄嵶嬠偺怤擖偲偼娭學側偔婲偒傞偙偲偑偁傝傑偡丅椺偊偽丄旍枮偼帀朾嵶朎傊偺帀幙偺拁愊偱偡偑丄帀朾慻怐撪偵儅僋儘僼傽乕僕偑怹弫偡傞偙偲偵傛傝帀朾慻怐撪偵枬惈揑側墛徢傪堷偒婲偟丄帀朾慻怐帺懱偑堎強揑側僒僀僩僇僀儞嶻惗偺応偲側傝傑偡丅傑偨丄摦柆峝壔偱偼寣娗撪暻偵晅拝偟偨僐儗僗僥儘乕儖摍偺帀朾惈捑拝暔幙偵儅僋儘僼傽乕僕偑堷偒婑偣傜傟偰丄偦偺儅僋儘僼傽乕僕偐傜曻弌偝傟傞僒僀僩僇僀儞偵傛偭偰嬊強揑偵敀寣媴嵶朎孮偑憹堳偟捑拝偟偨寢壥丄寣娗暻偺旍岤傪惗偠傑偡丅偡側傢偪丄旍枮偵偟傠摦柆峝壔偵偟傠丄枬惈揑側墛徢忬懺傪堎強揑偵惗傒弌偟偰偄傞偺偱偡丅偙偺傛偆側尰徾偼丄惗懱杊屼偲偄偆崌栚揑側壗傜偐偺棟桼偑偁傞偵偣傛丄寢壥偲偟偰惗懱偵埆塭嬁傪媦傏偡壜擻惈傪崅傔傑偡丅偮傑傝丄墛徢傪嬊強揑側僒僀僩僇僀儞偺棳擖偲偲傜偊偨応崌丄偦偺僒僀僩僇僀儞偵傛偭偰堷偒婲偝傟傞嵶朎墳摎偑丄偍偦傜偔廃埻偺嵶朎偵塭嬁傪梌偊傞偙偲偱丄娻偺敪徢傗恑峴偺儕僗僋傪崅傔偨傝丄條乆側戙幱幘姵傪堷偒婲偙偡壜擻惈偑偁傞偺偱偡乮恾俆乯丅 巹払偼丄偙偺傛偆側僸僩偵偍偗傞枬惈墛徢傗娻偺敪徢偲嵶朎惂屼偺攋抅偺娭楢傪尋媶偡傞偨傔偵丄庡偲偟偰儅僂僗屄懱傪梡偄偰夝愅傪峴偭偰偄傑偡丅慜弎偟偨傛偆偵丄巹払偼敪惗夁掱偵偍偄偰廳梫側栶妱傪壥偨偡Wnt僔僌僫儖揱払宱楬偵庡娤傪抲偄偰偙傟傑偱尋媶傪峴偭偰偒傑偟偨丅偟偐偟丄惉懱偵偍偄偰Wnt僔僌僫儖偺攋抅偑擛壗偵偟偰昦懺傗幘姵偲娭楢偡傞偐偵偮偄偰偼晄柧側晹暘偑懡偔懚嵼偟傑偡丅巹払偼偡偱偵婔偮偐偺Wnt娭楢場巕偺堚揱巕夵曄儅僂僗傪梡偄偰枬惈墛徢傗戙幱幘姵傪堷偒婲偡偙偲偱丄Wnt僔僌僫儖偑娭梌偡傞昦懺偺敪徢婡峔傪柧傜偐偵偟傛偆偲偟偰偄傑偡乮恾俇乯丅偙偺傛偆偵丄娻偺尋媶偵壛偊偰墛徢惈幘姵傗戙幱幘姵傪帇栰偵抲偄偰丄偙傟傜傪暘巕惗暔妛丒惗壔妛揑庤朄丄慻怐丒嵶朎妛揑庤朄丄堚揱巕夵曄丒昦懺儌僨儖儅僂僗傪梡偄偰摑崌揑偵夝愅偟棟夝偡傞偙偲傪栚巜偟偰偄傑偡丅巹払偼丄惗柦偺恀棟傗幘姵偺昦場傪棟夝偡傞偙偲偵尋媶偺幉懌傪抲偒傑偡偑丄巹払偺尋媶惉壥傪幘昦偺恌抐傗帯椕偵寢傃偮偗傞偙偲傪朰傟偢偵丄尋媶傪峴偭偰偄偒偨偄偲峫偊偰偄傑偡丅 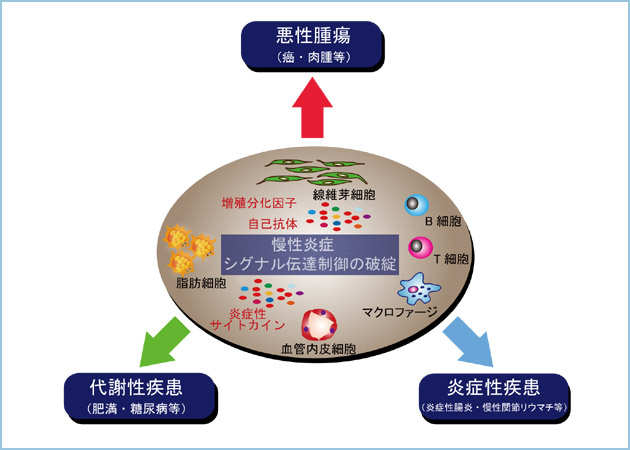 恾俆丏僔僌僫儖揱払惂屼偺攋抅傗枬惈墛徢偑娭梌偡傞幘姵丒昦懺
嵶朎傪庢傝姫偔娐嫬梫場偲偟偰枬惈墛徢傗僔僌僫儖揱払惂屼偺攋抅偼丄廃埻偺嵶朎偵庬乆偺埆塭嬁傪媦傏偡偙偲偱條乆側戙幱幘姵傪堷偒婲偡尨場偲側傞丅
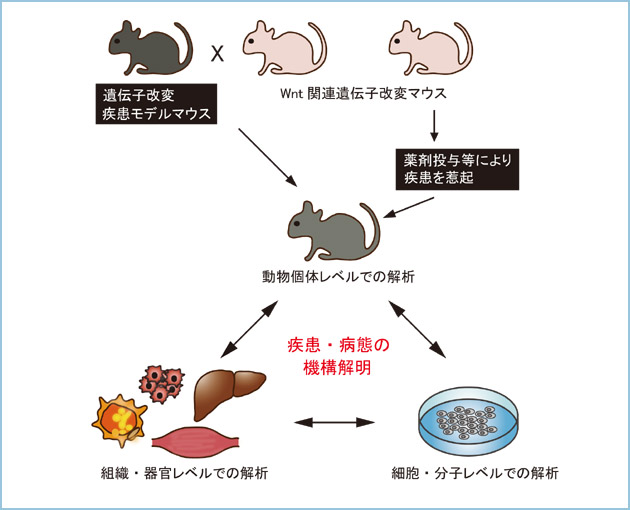 恾俇丏堚揱巕夵曄儅僂僗傪梡偄偨幚尡曽朄偺奣棯
Wnt娭楢場巕偺堚揱巕夵曄儅僂僗偵枬惈墛徢傗戙幱幘姵傪堷偒婲偡偙偲偱丄Wnt僔僌僫儖偑娭梌偡傞昦懺偺敪徢婡峔傪屄懱儗儀儖丒慻怐儗儀儖丒暘巕儗儀儖偱摑崌揑偵夝愅偟棟夝偡傞丅
偙偺傛偆側巹払偺尋媶巔惃偵嫟姶偟偰偄偨偩偗傞妛惗傗尋媶幰偺曽乆偼惀旕堦搙丄尋媶幒偵偍墇偟偔偩偝偄丅懡偔偺曽乆偺巹払偺尋媶傊偺嶲壛傪怱偐傜偍懸偪偟偰偄傑偡丅
|
|
| 戝嶃戝妛戝妛堾堛妛宯尋媶壢丂暘巕昦懺惗壔妛 | ||||||||||
|
 |