
現在の研究内容
1.電位センサーの動作原理
神経が興奮を伝達する仕組みは、長い間、生理学の最重要テーマのひとつとして研究されてきました。20世紀半ばに、ホジキンとハックスレーによりヤリイカの神経でナトリウムイオンが活動電位の形成に本質的な役割を担うことが発見され(Na説)、イオン電流の定量的な計測によって、(1)NaイオンとKイオンが別々の通路を通ること、(2)その通路の開閉は、膜電位と時間に依存して変化することが明らかにされ、電位依存性イオンチャネルの概念が確立しました。その後、様々な研究者により、トキシンへの結合を利用したイオンチャネルの単離や、パッチクランプ法による単一イオンチャネルを流れるpAレベルの微小電流の計測、分子クローニングによって電位依存性イオンチャネルの分子実体が明らかにされました。21世紀に入って、X線結晶構造解析によって、電位依存性カリウムチャネルなどの構造が原子レベルで明らかになるに到って、現在も最先端の研究が続いています。
私たちは、2005年と2006年に、世界に先駆けて、電位依存性イオンチャネルよりもシンプルな構造を有する、2つのユニークな膜蛋白質分子を発見しました(Murata et al, Nature, 2005; Sasaki et al, Science, 2006)。VSP(Voltage-Sensing Phosphatase)は、単一の電位センサーと、単一の酵素が分子内で連結した構造をしていて、細胞膜の電位が脱分極側へ上がると電位センサーの構造が変化し、その情報が細胞内の酵素領域に伝わり、ホスファターゼ(脱リン酸化酵素)の活性が出現します。つまり、電気シグナルが化学シグナルに変換されることになります。このホスファターゼ活性は、イノシトールリン脂質(リン脂質の一種で細胞内の様々な信号の制御に関わる)を脱リン酸化します。イノシトールリン脂質は、イノシトール環と呼ばれる糖の構造に共有結合しているリン酸の位置と数によって、そのシグナル分子としての役割が変わるため、この脱リン酸化によって細胞内の情報伝達が変化すると考えられます。
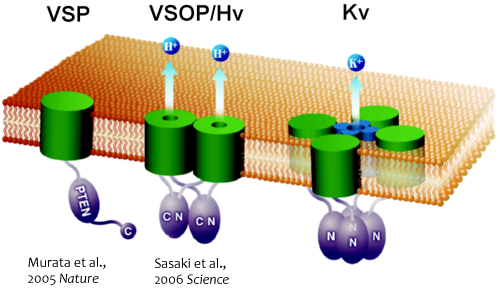
Fig.1 新規電位センサー蛋白質の発見
VSP : 電位依存性脱リン酸化酵素, VSOP : 電位依存性プロトンチャネル, Kv : 電位依存性カリウムイオンチャネル
一方、電位センサードメインのみから成るVSOP(Voltage sensor only protein)は、通常のイオンチャネルに見られるイオン透過ドメインをもっていないにも関わらず、水素イオンを通す働きのあるイオンチャネル(電位依存性プロトンチャネル:Hvチャネル)として機能します。膜電位を感知する機能しか持たないはずの電位センサードメインが、VSOPにおいては、膜電位感知と、プロトン透過の、一人二役の性質をもっています。この分子は、細胞内の領域を短く削っても(アミノ酸の長さにして100程度)イオンチャネル分子として機能できることから、哺乳類のゲノムでコードされているすべてのイオンチャネルのうち、最小で機能するイオンチャネルと言えます。
現在、VSPにおいてはどのように電位センサーの働きにより酵素が活性化するのか、VSOP/Hv1においてはどのように膜電位に応じてプロトン透過が起こるのかを、電気生理学的手法、光学的手法、構造生物学の手法を駆使して統合的に調べています。
2.ホスファターゼの動作原理
先に述べたVSPは、ガン化や代謝異常の原因として世界中で盛んに研究されている酵素PTENやチロシンホスファターゼと類似の構造を示すことから、VSPにおいて酵素活性が膜電位センサーにより制御される仕組みが明らかにされれば、これらの重要な酵素群の動作原理の解明につながる可能性があります。
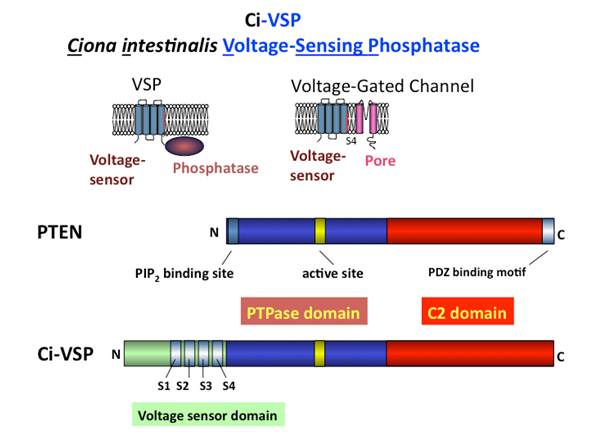
Fig.3 VSPとPTENの配列比較
脳腫瘍などの原因として重要な酵素であるPTENの酵素ドメインとVSPを比較して解析を行い、イノシトールリン脂質を基質として認識する分子基盤を明らかにしてきました(Iwasaki et al.,PNAS, 2008; Matsuda et al, JBC, 2011)。また、膜電位のレベルによって基質特異性が変化することを見出し、酵素の常識を超えるようなユニークな特性があることもわかりました(Kurokawa et al, PNAS, 2012)。これらの理解を深めることで、PTENやチロシンホスファターゼの酵素活性の制御機構の理解や創薬の分子ツールの確立につながり、ガン化や細胞内シグナル伝達の分野に貢献できるのでは、と考えています。また、ほとんどのタンパク質でも、機能ユニット(モジュール)が組み合わさることでタンパク質の機能が生み出されているので、VSPにおいて、電位センサーと酵素領域が共役する仕組みを明らかにすることで、タンパク質内でどのように機能ユニットが組み合わさるとタンパク質の機能が成立するのか、というタンパク科学全般に有用な研究になると考えています。
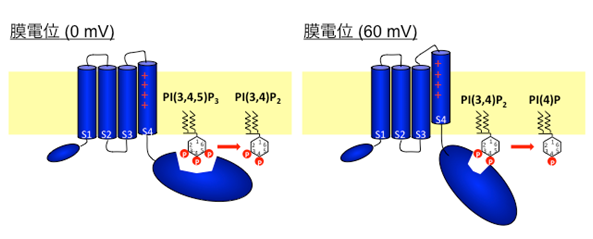
Fig.4 VSPのはたらき
3.血液細胞における電位依存性プロトンチャネルの役割
VSOP/Hv1(電位依存性プロトンチャネル=Hvチャネル)は海産無脊椎動物からヒトに到る様々な生物種に保存されており、哺乳類では、マクロファージや好中球といった貪食機能をもつ血球細胞に発現しています。プロトンを輸送する蛋白には、ATPを加水分解して細胞内からプロトンを運び出すプロトンポンプ(V-ATPase)がよく知られていますが、Hvチャネルは、プロトンポンプよりも短時間で大量のプロトンを排出することが可能です。膜電位の脱分極と細胞内外のpH差が引き金になってプロトン透過経路が開き、これによりプロトンイオン濃度勾配に従ってプロトンが通過するので、直接ATPを必要としません。
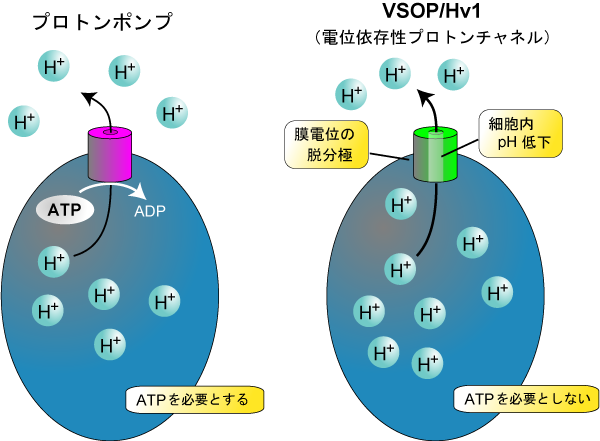
Fig.5 電位依存性プロトンチャネル(VSOP/Hv1)の役割
これまでの電気生理学的な研究から、Hvチャネルは、貪食細胞での活性酸素の産生を促進することが示唆されてきました。最初に貪食細胞で活性酸素が作られる際には、酸素がO2-というマイナスの電荷をもつ物質として細胞の外へ放出されます。この状態が続くと電荷のアンバランスのため、細胞膜が大きく脱分極してしまい、オキシダーゼの活性が減弱してしまいO2-の産生が止まってしまいます。この電荷のアンバランスをキャンセルする役者としてプロトンを輸送する経路が想定され、Hvチャネルはまさにこの経路を担う実体として研究されてきました。私たちは、このHvチャネルの分子実体の同定に成功し(Sasaki et al, Science, 2006)、更にVSOP/Hv1のノックアウトマウスを用いた解析を行い、プロトンの輸送により細胞質のpHと膜電位を制御することで、活性酸素の産生を助ける働きがあることを明らかにしました(Okochi et al, BBRC, 2009, El Chemaly et al, JEM, 2010、スイスのジュネーブ大学との共同研究)。現在、pHイメージング、膜電位計測、などを行って、これまで未知であった生体レベルでの電位依存性プロトンチャネルの役割を明らかにしつつあります。
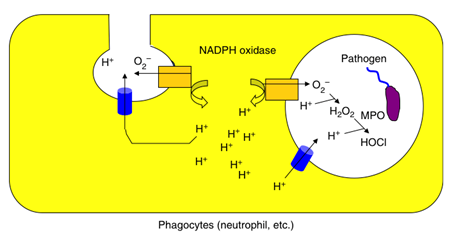
Fig.6 電位依存性プロトンチャネル(VSOP/Hv1)の生体内での役割

Fig.7 電位依存性プロトンチャネル(VSOP/Hv1)の配列比較
4.膜電位の可視化にむけた分子ツール開発
神経回路の機能を理解する上で、個々のニューロンの活動を同時に計測することは極めて重要であり、また、電気生理学は、ガラス電極などが物理的にアプローチできない細胞や組織には適用することは困難で、膜電位の可視化は、こうした細胞での電気的現象の解明に有用であると考えられます。これまで有機化合物を用いた電位感受性色素やCa感受性色素がこれらの解析に用いられてきています。しかし、特定の細胞のみにプローブをターゲットさせることが困難であったり、シグナルの強度が強くないなどの問題があり、普遍的に用いられる手法は、いままで確立されていませんでした。最近、複数のグループによりVSPの電位センサーに蛍光蛋白を結合させた分子が作成されました。我々の研究室のメンバーである筒井助教は、理化学研究所の宮脇博士のグループに在籍時に、生理学研究所と協力して、蛍光強度変化の大きいプローブ(海産動物由来のタンパクを組み合わせたことから、Mermaidと命名)を作り出すことに成功し、培養細胞レベルではありますが、ニューロンや心筋などの電気活動をイメージング化することに成功しました(Tsutsui et al., Nature Methods, 2008)。現在、私たちはこれらの膜電位プローブ分子の改良を行い、将来的にはin vivoの組織での計測に応用する技術を目指しています。
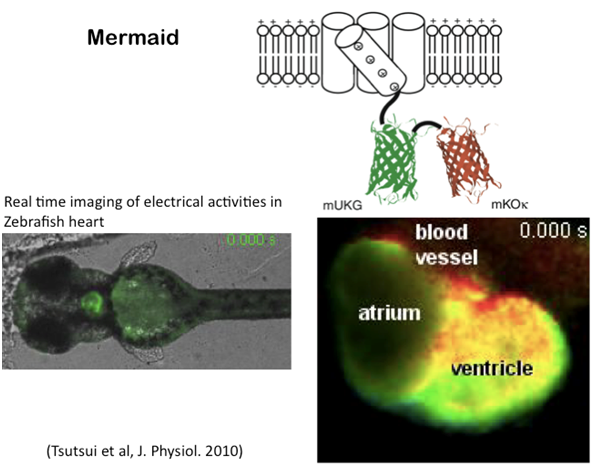
Fig.8 膜電位可視化へ向けてのツール開発
5.生命現象における、新たな膜電位変化の役割の解明
電位センサー蛋白の発見により膜電位を伝える全く新しい分子経路が明らかとなり、これまでの膜電位の生理機能の役割を見直す必要が出てきました。たとえば、受精における膜電位シグナルによる多精子拒否現象や、非哺乳類脊椎動物での左右軸決定過程や再生過程で、細胞膜電位変化の関与が示唆されてきたものの、未だに具体的な分子機構は明らかになっていません。現在複数の共同研究者とともに、VSPの生理的な役割を調べる研究を行っています。また、筒井らが開発したMermaidを用いて、これまで膜電位変化が調べられてこなかった現象で、膜電位変化が起こっているかを調べようとしています。これらにより、再生や発生過程などでの膜電位シグナルの新たな役割が明らかになるのではと期待しています。
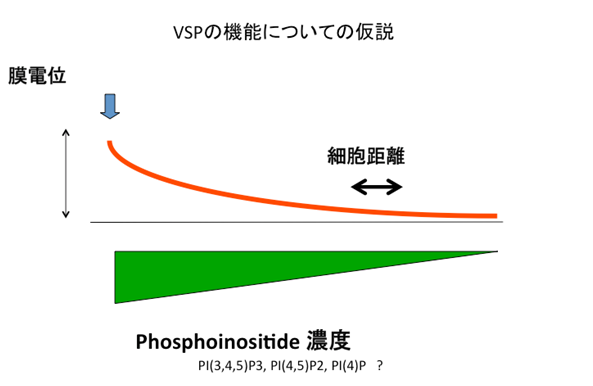
Fig.9 VSPの生理機能
VSPはゲノム情報から発見され、膜電位に依存して酵素活性が変化する分子機能をもつ分子であるが、その実際の生理機能は良くわかっていない。精巣などに発現することから、精子の機能や、発達に重要な役割を果たしている可能性がある。図は、分子特性から想定できるメカニズムを示している。もしVSPタンパクが細胞膜上に均一に存在するならば、膜電位の勾配に応じて細胞内でのイノシトールリン脂質の濃度が変化すると予測される。(adopted from “Biodiversity of voltage sensor domain proteins”by Y. Okamura,Pflugers Archiv., 2007)
6.人工網膜を中心とした視覚機能回復の基礎的研究(三好智満)
光を電気信号に変換する網膜細胞(視細胞)が変性する網膜色素変性や加齢黄斑変性などの疾患では、視覚は永続的に失われ、その治療法はまだありません。このような失明者の視機能を回復させる1つの手段として人工網膜があり、世界中で研究・開発が進んでいます。多くの人工網膜では、網膜に多数の電極を貼り付けて電流を流し、残っている網膜神経回路を直接興奮させることによって光の点(擬似光覚)で構成された視覚像を発生させます。我々は網膜への侵襲を避けるため、強膜上に置いた多点電極と硝子体内に設置した帰還電極間で通電する脈絡膜上-経網膜刺激 suprachoroidal-transretinal electrical stimulation (STS)方式の人工網膜を考案し、その有効性を報告しました(Kanda, et al., Invest Opthal Vis Sci., 2005)。その後も電気生理学的な方法や光学計測法を用いながら、STSによる網膜興奮のメカニズムを明らかにし、より効率的で安全な人工網膜システムの仕様を探る基礎研究を行っています。2010年にはSTS方式の人工視覚システムを使った亜急性臨床試験が本学感覚機能形成外科学・眼科学のグループ(代表:不二門尚教授)により行われ、試験に協力して頂いた二名の網膜色素変性の患者さんの眼に装着したシステムで、物体の位置と大きさの大まかな区別が可能になることが判りました。また、我々は網膜神経細胞に対する電気刺激の保護効果を見いだし、その効果を付与した人工網膜システムの開発に取り組んでいます。この人工網膜の研究・開発には、医学、工学など複数の分野間の緊密な連携が重要なので、上述の眼科学・感覚機能形成学講座だけでなく、本学工学研究科(八木哲也教授)、奈良先端科学技術大学(太田淳教授)、株式会社ニデック人工視覚研究所と連携して研究を進めています。