Wntシグナルの選択的活性化
受容体エンドサイトーシスを介する選択的活性化
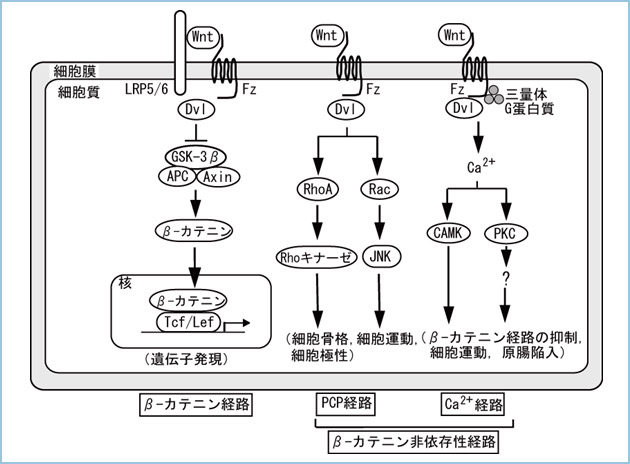
Wntが細胞に作用することにより、活性化される細胞内シグナル伝達機構をWntシグナル経路と呼びますが、本シグナル経路には①β-カテニンを介して遺伝子発現を制御するβ-カテニン経路、②細胞の平面内極性を制御するPCP(planar cell polarity, 平面内細胞極性)経路、③Ca2+の細胞内動員を促進するCa2+経路の少なくとも3種類が存在します(図1)。Wntシグナル伝達機構の中で最もよく知られているのが、β-カテニン経路です。β-カテニンはカドヘリン結合蛋白質として同定され、細胞接着に重要な働きをしますが、Wntシグナルのメディエーターとして遺伝子発現を誘導し、その結果、β-カテニン経路は細胞の増殖や分化を制御することが明らかになっています。一方、Wntシグナルにはβ-カテニンに依存しない細胞内シグナル伝達機構が少なくとも2経路存在します(β-カテニン非依存性経路)。第一の経路であるPCP経路は、ショウジョウバエの翅毛の配向を決定するシグナルとして見出されました。PCP経路はWnt受容体Fz (Frizzled) と細胞内蛋白質Dsh/Dvl (Dishevelled) を介して、Rhoファミリーの低分子量G蛋白質(GTP結合蛋白質)を活性化し、さらにRho-キナーゼやJNK (Jun-N-terminal kinase, Junキナーゼ) の活性化により、細胞骨格や細胞運動、遺伝子発現を調節します。第二の経路であるCa2+経路は、Wntが細胞内Ca2+を動員し、CaMK (カルモデュリン依存性蛋白質リン酸化酵素) とPKC (蛋白質リン酸化酵素C) を活性化します。この2つの経路に加えて、新たな経路も報告されつつあります。
図1.Wntシグナル経路の多様性
Wntシグナルはβ-カテニン経路、PCP経路、Ca2+経路の3つの経路を活性化する。
CaMK; Ca2+/calmodulin-dependent kinase, Fz; Frizzled, GSK-3β; glycogen synthase kinase-3β, JNK; Jun N-terminal kinase, LRP5/6; low-density-lipoprotein receptor-related protein 5/6, PKC; Protein kinase C, Tcf/Lef; T-cell factor/lymphoid enhancer factor.
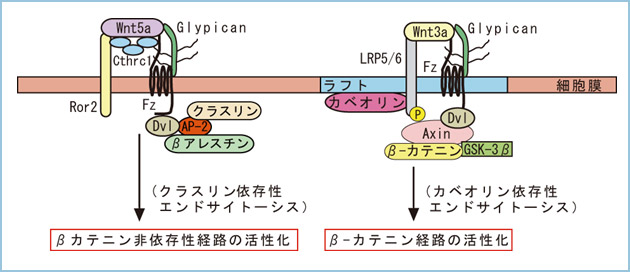
Wnt遺伝子はヒトやマウスのゲノム上に19種類存在し、大きなファミリーを形成しています。また、Wnt受容体として7回膜貫通型Frizzledが10種類、その共役受容体として、1回膜貫通型LRP5, LRP6, Ror1, Ror2, Ryk等が存在します。大きな疑問はこの多様なWntと受容体の組み合わせが、どのようにして上述した複数のシグナル経路を選択的に活性化するかということです。共役受容体のLRP5/6はβ-カテニン経路の活性化に関与し、Ror1/2はβ-カテニン非依存性経路の活性化に関与することが明らかになっています。私達はWntと受容体の特異的な組合せに加えて、受容体エンドサイトーシス経路が重要であると考えています。例えば、Wnt3a依存性のβ-カテニン経路の活性化(β-カテニンの安定化)にはカベオリン依存性のエンドサイトーシス経路が重要ということがわかってきました。また、細胞膜上のリピッドラフトの存在も重要です。しかし、Wnt5a依存性のβ-カテニン非依存性経路の活性化(Racの活性化)にはクラスリン依存性のエンドサイトーシス経路が関与していました。本経路の活性化には、リピッドラフトの存在は関係しませんでした。このように、細胞膜上のマイクロドメインとそれに関連する受容体エンドサイトーシス経路がWntシグナルの選択的活性化を制御すると考えています(図2)。
図2.細胞膜上でのWntシグナルの選択的活性化の制御機構
WntはFz受容体と共役受容体LRP5/6やRor2に加えて、Glypican等の膜結合型糖蛋白質やCthrc1等の分泌型糖蛋白質との組み合わせによってβ-カテニン経路とβ-カテニン非依存性経路を選択的に活性化すると考えられる。私達は、Wnt3aによってLRP6/Fz5受容体がカベオリン依存性エンドサイトーシスされることがβ-カテニンの安定化に必要であることを示している。また、β-カテニン非依存性経路の活性化にはクラスリン依存性エンドサイトーシスが関与することも明らかにしている。
ヘパラン硫酸プロテオグリカンを介する選択的活性化
Wntシグナルを制御する因子の一つとしてプロテオグリカンがあります。プロテオグリカンは核となる蛋白質に多数のグリコサミノグリカン鎖(糖鎖)が結合しています。プロテオグリカンは様々な成長因子と結合して、その空間的配置や機能を制御することによって多様な細胞機能に関与すると考えられています。プロテオグリカンの一つであるGlypican(GPC)はGPIアンカーによって細胞表面に結合しており、Wntシグナルに関与することが報告されています。
上述したように、私達はこれまでにWntβ-カテニン経路の活性化時に起きる受容体LRP6のリン酸化やカベオリン依存性エンドサイトーシスは、脂質ラフトと呼ばれるシグナル伝達の足場となるマイクロドメインで起きることを明らかとしています。GPCは脂質ラフトに局在し、Wntと結合することができます (図1)。Glypican4 (GPC4) はこれらの性質によって、脂質ラフトにWntを集積することでβ-カテニン経路がより活性化しやすい環境を作り出していることが明らかになりました。また、GPC4は非脂質ラフトにも局在し、このGPC4はβ-カテニン非依存性経路を活性化することがわかりました。
図1.細胞膜上でのWntシグナルの選択的活性化の制御機構
WntはFz受容体と共役受容体LRP5/6やRor2に加えて、Glypican等の膜結合型糖蛋白質やCthrc1等の分泌型糖蛋白質との組み合わせによってβ-カテニン経路とβ-カテニン非依存性経路を選択的に活性化すると考えられる。私達は、Wnt3aによってLRP6/Fz5受容体がカベオリン依存性エンドサイトーシスされることがβ-カテニンの安定化に必要であることを示している。また、β-カテニン非依存性経路の活性化にはクラスリン依存性エンドサイトーシスが関与することも明らかにしている。
GlypicanはGPIアンカーの部分がリパーゼによって切断されることで、細胞膜から解離することが知られています(図2)。細胞膜から解離したGPC4 ectodomainはWntと結合する性質は維持されていますが、細胞膜上のGPC4と異なり、β-カテニン経路とβ-カテニン非依存性経路を抑制しました。細胞膜から解離したGPC4 ectodomainはWnt3aやWnt5aとそれらの受容体との結合を阻害しました。
このようにGlypicanがWntシグナルを制御するには脂質ラフトと非脂質ラフトに局在すること、また、細胞膜にアンカーしているか、あるいは細胞膜から解離しているか等の性質が重要であり、Glypicanのこれらの性質がどのようにコントロールされているのかを解析することが今後重要になってくると考えています。
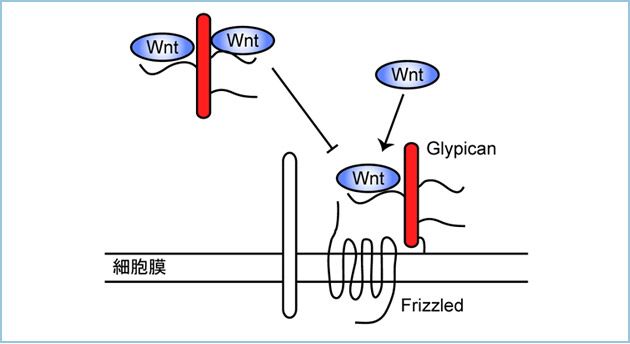
図2.細胞膜から解離したGlypicanによるWntシグナルの制御
GlypicanはGPIアンカーがリパーゼにより切断されることで細胞膜から解離する。細胞膜から解離したGlypicanはWntと受容体の結合を阻害する。