上皮細胞におけるWntの極性分泌制御
肝細胞におけるLRP6の異なるエンドサイトーシス経路の制御
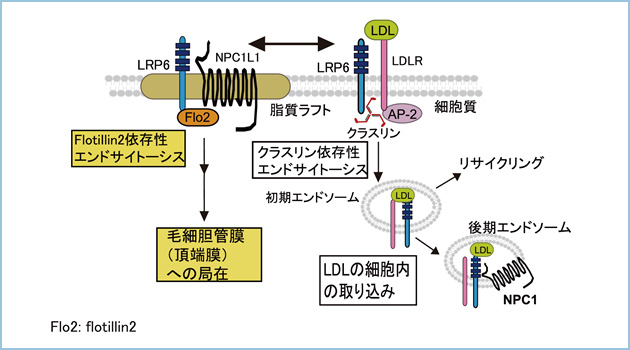
私達は、Wnt3aとDKK1がLRP6をそれぞれカベオリン依存性とクラスリン依存性経路でエンドサイトーシスさせることにより、Wntシグナルを活性化あるいは抑制することを明らかにしてきました。また、LRP6はlow-density lipoprotein(LDL)刺激によってLDL受容体と複合体を形成し、LDL受容体とLDLの細胞内輸送を促進することにより、脂質代謝制御にも関与することが示唆されました。したがいまして、LRP6は種々のリガンドに応答し、エンドサイトーシスされることにより細胞応答を制御しますが、脂質代謝におけるLRP6の輸送経路と機能は不明です。
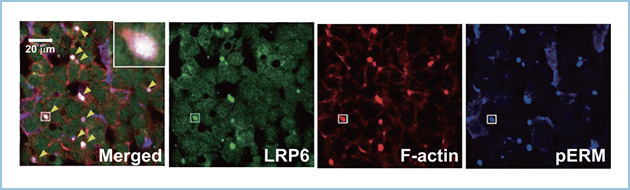
肝細胞は隣接する細胞間に密着結合が形成されることにより頂端側と側底側が区分され、頂端側には2個から数個の細胞に囲まれて胆汁の通路である毛細胆管が形成されます。肝細胞の側底側には類洞(血管)や血漿が含まれるディッセ腔が存在し、類洞からのタンパク質や血漿成分が側底膜から肝細胞内に取り込まれます。ヒト肝がん細胞株HepG2細胞を4-5日培養しますと密着結合が形成され、頂底極性を獲得し、頂端側に毛細胆管様構造が形成されます。そこで、HepG2細胞を用いて脂質代謝におけるLRP6の輸送経路と機能を解析しました。最初に内在性のLRP6の局在を解析したところ、LRP6は頂端膜に局在しました(図1)。
図1.肝細胞におけるLRP6の局在
HepG2細胞において内在性のLRP6は頂端膜マーカーのF-アクチンやリン酸化ERM(pERM)と頂端膜(黄色矢頭)において共局在した。
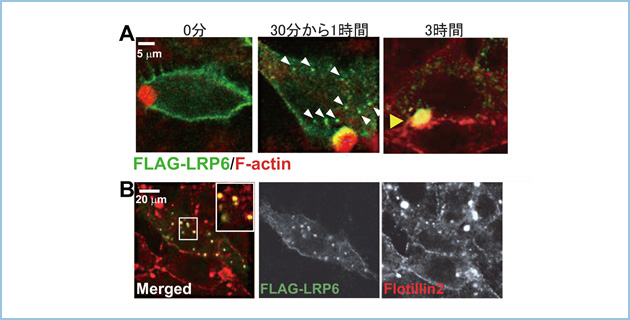
HepG2細胞にFALG-LRP6を発現させ、側底膜に局在するFLAG-LRP6をFLAG抗体により標識した後、FLAG-LRP6の局在を経時的に解析しました。その結果、側底膜に局在するFLAG-LRP6は30分〜1時間後に細胞質に移行し、3時間後には頂端膜に輸送されました(図2A)。また、エンドサイトーシスされたLRP6は頂端膜への輸送を制御するflotillin2と共局在しました(図2B)。この結果から転写・翻訳されたLRP6はトランスゴルジから一度側底膜に輸送された後、flotillin2依存性に頂端膜へ輸送(トランスサイトーシス)されることが示唆されました。
図2.肝細胞におけるLRP6の輸送経路
A. HepG2細胞にFALG-LRP6を発現させ、4℃においてFLAG抗体を含むメディウムとインキュベートすることにより側底膜に局在するFLAG-LRP6を一度FLAG抗体により標識した後、FLAG-LRP6の局在を経時的に解析した。側底膜に局在するFLAG-LRP6は37℃で培養した30分〜1時間後に細胞質(白色矢頭)に移行し、3時間後に頂端膜(黄色矢頭)に局在した。
B. 側底膜に局在するFLAG-LRP6をFLAG抗体により標識し、37℃で1時間培養した後、FLAG-LRP6の局在を解析したところ、flotillin2と共局在した。
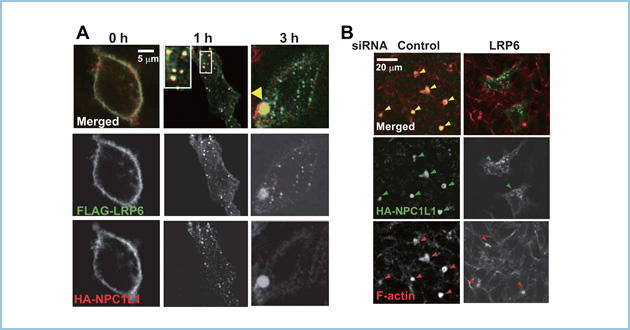
次にLRP6が肝細胞の頂端膜に局在する意義を検討しました。肝細胞において、Niemann Pick C1-like 1 (NPC1L1)は頂端膜(毛細胆管膜)に局在し、コレステロールの吸収に関与することが知られています。私達は定常状態ではNPC1L1がLRP6と挙動を共にし、細胞膜から細胞質へ移行し、頂端膜で共局在することを見出しました(図3A)。また、NPC1L1の頂端膜への局在はLRP6に依存することを見出しました(図3B)。これらの結果から、NPC1L1はLRP6と結合することにより、側底膜から頂端膜へトランスサイトーシスすることが示唆されました。
図3.HepG2細胞におけるNPC1L1の頂端膜へのトランスサイトーシス
A. HepG2細胞にFLAG-LRP6とHA-NPC1L1を発現させ、側底膜に局在するFLAG-LRP6とHA-NPC1L1をFLAG抗体とHA抗体により標識した後、両者の局在を経時的に解析した。37℃で培養した1時間後に両者は細胞質において共局在し、3時間後には頂端膜(黄色矢頭)に局在した。
B. HepG2細胞にHA-NPC1L1を発現させ、側底膜に局在するHA-NPC1L1をHA抗体で標識した後、37℃で培養して3時間後の局在を解析した。LDL非存在下(コントロール)ではHA-NPC1L1は頂端膜に局在したが(黄色矢頭)、LRP6を発現抑制するとHA-NPC1L1は頂端膜(F-actin、赤色矢頭)へのトランスサイトーシスが抑制され、細胞質に局在した。
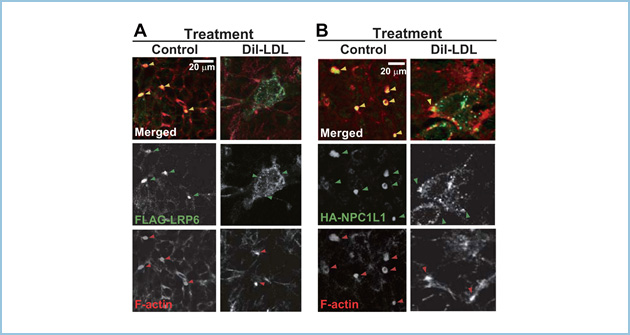
一方、HepG2細胞をLDLで刺激した場合、LRP6はクラスリン依存性エンドサイトーシスされることにより、頂端膜へのトランスサイトーシスが阻害されました。同様に、LDL刺激によりNPC1L1の頂端膜への局在が抑制されました(図4A,B)。
図4.LDL刺激によるLRP6とNPC1L1の頂端膜へのトランスサイトーシスに対する影響
A, B. FLAG抗体とHA抗体により、FLAG-LRP6とHA-NPC1L1を標識した後、Dil-LDL存在下と非存在下において37℃で3時間培養した後、FLAG-LRP6(A)とHA-NPC1L1(B)の局在を解析した。Dil-LDL非存在下ではFLAG-LRP6とHA-NPC1L1は頂端膜に局在したが(黄色矢頭)、Dil-LDL刺激により、FLAG-LRP6とHA-NPC1L1は頂端膜(F-actin、赤矢頭)へのトランスサイトーシスが抑制され、細胞質に局在した。
したがいまして、LDLの有無によって肝細胞におけるLRP6のエンドサイトーシスの経路が変化し、それに伴ってNPC1L1の頂端膜への局在が制御されることが示唆されました。
これらの結果から、定常状態ではLRP6がNPC1L1と複合体を形成し、NPC1L1を毛細胆管膜に輸送することにより、胆汁からコレステロールの吸収を促進すると考えられます。一方、LDL存在下ではLRP6はLDL受容体と共にクラスリン依存性エンドサイトーシスによって細胞質に取り込まれます。その結果、LRP6によるNPC1L1の毛細胆管膜への輸送を抑制することで、胆汁からのコレステロールの吸収を調節していると考えられます(図5)。
図5.肝細胞におけるLDLによるLRP6とNPC1L1の局在制御
肝細胞において、LDLの有無によってLRP6のエンドサイトーシス経路が決定され、NPC1L1の頂端膜への局在が制御される。LDL非存在下において側底膜に局在するLRP6はNPC1L1と共にflotillin2依存性エンドサイトーシスにより細胞質に移行した後、頂端膜へ輸送される。LDL存在下ではLRP6はLDL受容体と共にクラスリン依存性エンドサイトーシス経路により細胞質に移行することにより、LDLを細胞内に取り込む。その結果、NPC1L1の頂端膜への移行が抑制される。
Wntの脂質修飾と分泌
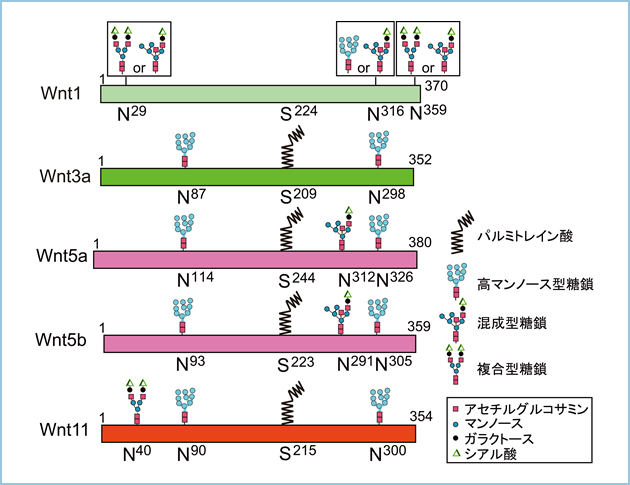
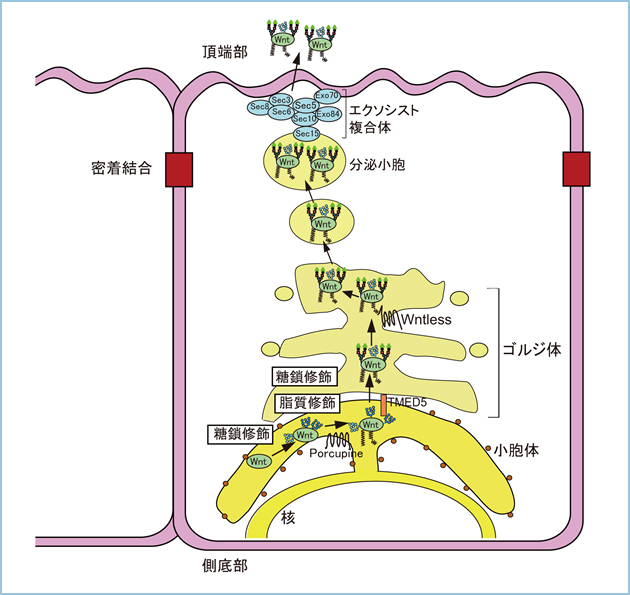
Wntは小胞体やゴルジ体において脂質や糖鎖の翻訳後修飾を受け、機能発現が制御されています。Wntの脂質修飾についてはWnt3aについて解析が進められており、209番のセリンにパルミトレイン酸が付加されることが報告されています。この脂質修飾されるセリン残基はWnt1をはじめ、他のWntファミリーにも保存されており、Wnt3aと同様に脂質修飾されることが示唆されます。事実、私達はβ-カテニン非依存性経路を活性化する代表的なリガンドのWnt5aやWnt5b、Wnt11もパルミトレイン酸が付加されることを見出しました(図1)。Wntのパルミトレイン酸の付加は小胞体に局在するアシル転移酵素Porcupineによって制御されています。また、Wntの細胞外分泌を制御するWntlessとの結合にパルミトレイン酸の付加が必要であり、パルミトレイン酸が付加されないWnt3aやWnt5a、Wnt5b、Wnt11変異体はWntlessとの結合能が無く、細胞外に分泌されませんでした。したがいまして、Wntの脂質修飾は細胞外分泌に重要な機能を果たすと考えられます。
図1.Wntの翻訳後修飾
Wnt3aの209番、Wnt5aの244番、Wnt5bの223番、Wnt11の215番のSにはパルミトレイン酸が付加される。他のWntと同様にWnt1において224番にSが保存されており、パルミトレイン酸が付加されると考えられる。また、Wnt1の316番のNには高マンノース型か混成型、29番と359番のNには混成型か複合型糖鎖が付加される。Wnt3aの87番と298番のNには高マンノース型のN結合型糖鎖が付加される。Wnt5aの114番と326番のNには高マンノース型、312番のNには混成型のN結合型糖鎖が付加され、同様にWnt5bの93番と305番のNには高マンノース型、N291番のNには混成型のN結合型糖鎖が付加される。一方、Wnt11の90番と300番のNには高マンノース型、40番のNには複合型のN結合型糖鎖が付加される。
Wntの糖鎖修飾と分泌
Wntは繊維芽細胞や血球系細胞の間質細胞に加えて上皮細胞に発現しています。繊維芽細胞を用いた解析から、小胞体からゴルジ体への輸送にTMED5(transmembrane emp24 protein transport containing domain 5)、ゴルジ体から細胞外への輸送にはWntlessが関与することは報告されていますが、詳細な分泌機構は明らかにされていません。一方、極性化された上皮細胞において膜タンパク質や分泌タンパク質がトランスゴルジ網(TGN)から頂端部、あるいは側底部へ異なる機構で選別輸送されることが明らかになりつつあります。これまでに、Wntシグナルが細胞極性の制御に重要な機能を果たしていることは示されていますが、Wnt自身が極性化された上皮細胞において頂端部、側底部のいずれに分泌されるかは解析されていませんでした。また、Wntはアスパラギン(N)結合型糖鎖が付加されますが、Wntの糖鎖構造と細胞内輸送との関連は不明です。そこで、私達はWnt3aやWnt5a、Wnt11の糖鎖構造と上皮細胞における極性分泌との関連を解析しました。質量分析法により糖鎖構造を解析した結果、Wnt3aの87番と298番のNに高マンノース型、Wnt5aの114番と326番のNに高マンノース型、312番のNに混成型、Wnt11には90番と300番のNに高マンノース型、40番のNには複合型糖鎖が付加されていました。また、Peptide:N-glycosidase FやEndoglycosidase Hのグリコシダーゼを用いてWnt1の糖鎖修飾を解析した結果、Wnt1の316番のNに高マンノース型か混成型、29番と359番のNに混成型か複合型糖鎖が付加されていることが示唆されました(Wntの脂質修飾と分泌、図1)。
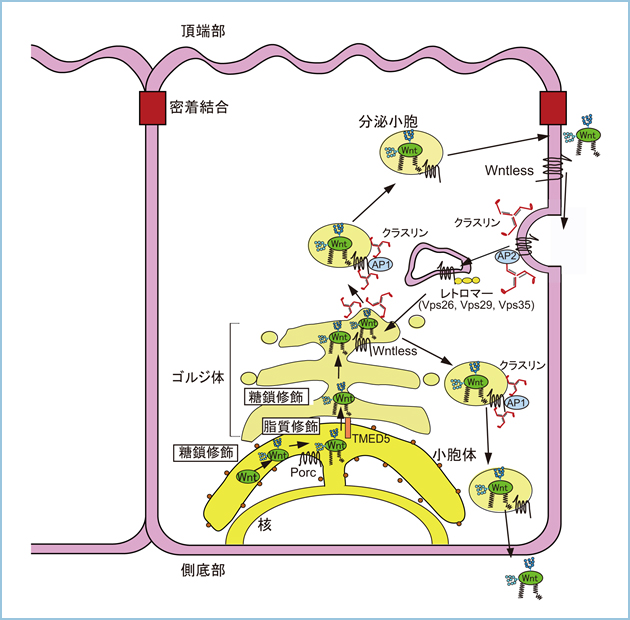
続いて、極性化されたイヌ腎臓上皮由来MDCK細胞においてWntの細胞外分泌を解析した結果、Wnt3aとWnt5aは側底部、Wnt11は頂端部、Wnt1は側底部と頂端部の両方向に分泌されることが明らかになりました(図1-3)。さらに、Wnt1やWnt3a、Wnt5aはWntlessと共にクラスリンとアダプター蛋白質(AP)複合体のAP-1を介してTGNから側底部へ選別輸送されました(図1)。これまでに、細胞膜上でWntを解離したWntlessはクラスリン依存性エンドサイトーシスを介して細胞質に移行した後、レトロマー複合体によってゴルジ体へリサイクルされることが報告されていましたが、Wnt3aの側底部への分泌にはこのWntlessのリサイクル経路が必要でした。一方、クラスリン、ならびにAP-1やAP-2の機能発現を抑制してもWnt11の頂端部への輸送には影響しないことから、Wnt11はWntlessとの複合体形成を介さず、TGNから頂端部へ極性分泌されることが示唆されました。
図1.極性化された上皮細胞におけるWnt1やWnt3a、Wnt5aの側底部への分泌制御
小胞体においてWnt3aとWnt5aは糖鎖修飾とPorcupineによるパルミトレイン酸の脂質修飾を受ける。TMED5によって小胞体からゴルジ体へ輸送されたWnt1やWnt3a、Wnt5aはTGNにおいてWntlessと共にクラスリン/AP-1を介して側底部へ選別輸送され、細胞外に分泌される。一方、細胞膜上でWntを解離したWntlessはクラスリン依存性エンドサイトーシスを介して細胞質に移行した後、レトロマー複合体によってゴルジ体へ輸送される。
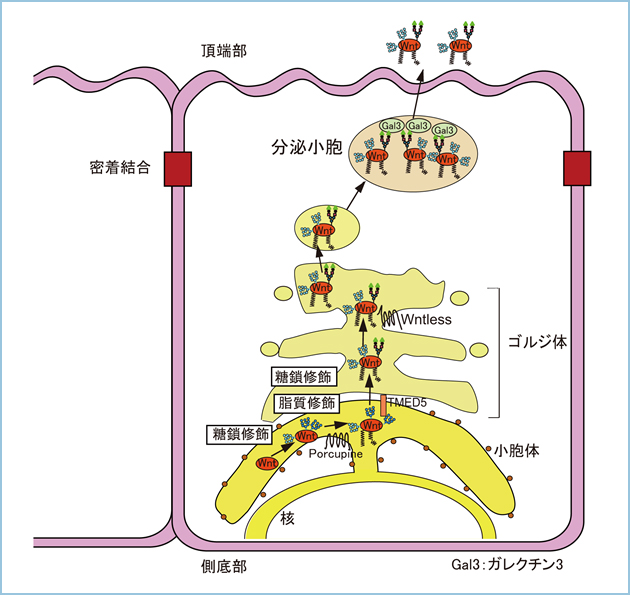
極性化された上皮細胞において膜タンパク質の頂端部への輸送シグナルの一つにN結合型糖鎖が報告されています。Wnt3aとWnt11の糖鎖修飾の違いを考えると、Wnt11のN末端側に付加される複合型糖鎖が極性分泌に関連すると考えられました。そこで、極生化されたMDCK細胞において複合型糖鎖修飾を抑制し、高マンノース型のみの糖鎖修飾にするとWnt11の頂端部への極性分泌が抑制されました。さらに、Wnt11のN40を含むN末端領域 (1-52アミノ酸)をWnt3aのC末端領域(50-352アミノ酸)に融合させたWnt11/3aのキメラタンパク質はWnt11と同様にN40には複合型糖鎖が付加され、その結果、Wnt11/3aは極性化されたMDCK細胞において頂端部へ分泌されました。同様にWnt3aやWnt5aのN末端側に糖鎖修飾部位を挿入したWnt3aL35NとWnt5aL68Nはいずれも複合型糖鎖が付加され、頂端部へ分泌されました。すなわち、Wnt11の極性分泌において、N末端側に複合型糖鎖が付加されると側底部よりも頂端部への輸送シグナルが優位に働き、Wntタンパク質が頂端部へ分泌されると考えられます(図2)。
これまでに、複合型糖鎖が付加されたタンパク質の細胞内輸送を制御する分子としてガラクトースと結合するガレクチンが報告されています。ガレクチンとWnt11の極性分泌の関連を解析したところ、ガレクチン3の発現を抑制するとWnt11の頂端部への極性分泌が抑制されました。したがいまして、TGNから遊離したWnt11を含む小胞はガレクチン3が含まれるエンドソームに融合した後、Wnt11がガレクチン3に結合することにより、頂端部へ分泌されると考えられます(図2)。
図2.極性化された上皮細胞におけるWnt11の頂端部への分泌制御
小胞体においてWnt11は糖鎖修飾とPorcupineによるパルミトレイン酸の脂質修飾を受ける。TMED5によって小胞体からゴルジ体へ輸送されたWnt11はさらにN結合型糖鎖のプロセシングを受け、40番のNの糖鎖は複合型に変換される。TGNにおいてWnt11はWntless非依存性に頂端部に選別輸送され、Wnt11が取り込まれた小胞はガレクチン3が存在する小胞に融合した後、細胞外に分泌される。
一方、Wnt1はN末端側の29番のNへの複合型糖鎖の付加を抑制しても頂端部への分泌は抑制されませんでしたが、Sec8やSec6を含むエクソシストを介して分泌されることを見出しました(図3)。したがって、Wntによって糖鎖修飾部位や付加される糖鎖構造に加えて、極性された上皮細胞におけるWntの分泌機構が異なることが明らかになりました。
図3.極性化された上皮細胞におけるWnt1の頂端部への分泌制御
小胞体からゴルジ体へ輸送されたWnt1はN結合型糖鎖のプロセシングを受け、混成型や複合型に変換される。Wnt1が取り込まれた小胞はSec8やSec6を含むエクソシストを介して頂端部へ選別輸送され、細胞外に分泌される。
Wnt受容体の極性輸送
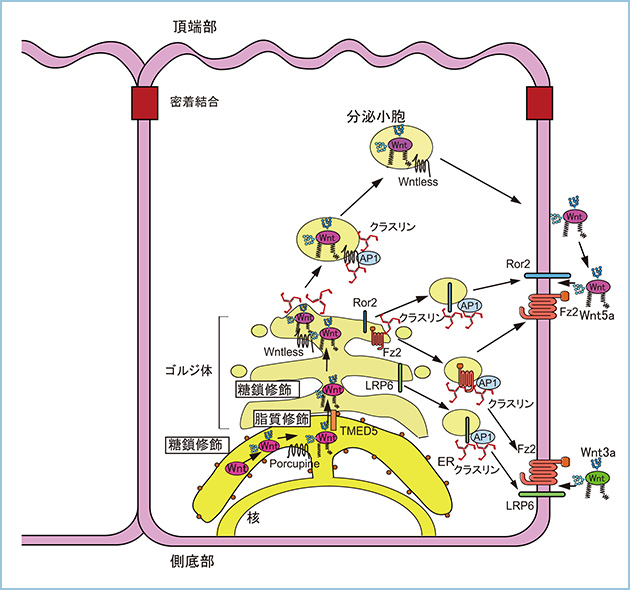
Wntシグナルの選択的活性化は7回膜貫通型受容体のFrizzled(Fz)と一回膜貫通型受容体のLRP6やRor2、Rykの組み合わせによって決定されます。LRP6はWnt3aに結合しβ-カテニン経路を活性化し、Ror2とRykはそれぞれWnt5aとWnt11に結合しβ-カテニン非依存性経路を活性化しますが、上皮細胞におけるWnt受容体の極性輸送については明らかにされていません。そこで、極性化されたMDCK細胞においてWnt受容体の輸送経路について解析しました。その結果、Fz2やLRP6、Ror2は側底側に局在し、Fz7やRykは頂端部、側底部の両者に局在しました。また、Wntlessの発現抑制によってWnt受容体の極性輸送には影響しませんが、クラスリンやAP-1の発現抑制によってFz2やRor2、LRP6の側底部への輸送が抑制されました(図1)。これはWntとその受容体は別々の小胞によって側底部へ輸送され、細胞膜への輸送時に両者の結合によるWntシグナル活性化が誘導されないためと考えられます。また、Wnt3aやWnt5aを頂端部から作用させてもWntシグナルは活性化されませんが、側底側から刺激するとWntシグナルが活性化されました。したがって、WntとWnt受容体が極性化上皮細胞の同じ領域に輸送されることにより、効率よくWntシグナルが活性化されると考えられます。
図1.上皮細胞におけるWnt受容体の極性輸送の制御
7回膜貫通型受容体のFz2とその共役受容体のLRP6やRor2はクラスリン/AP-1を介して側底部へ極性輸送される。一方、これらのWnt受容体はWntless非依存性に側底部へ極性輸送されることから、Wntとその受容体は別々の小胞によって側底部へ輸送される。Wnt リガンドとWnt受容体を同じ側底部に輸送することにより、効率よくWntシグナルが活性化される。