Wntシグナルによる細胞機能制御
Prickleを介した新規の細胞極性、運動の制御機構
私達はこれまでに、β-カテニン非依存性Wntシグナル経路を活性化するWnt5aに着目し、Wnt5aシグナルが細胞運動や接着を制御する仕組みを明らかにしてきました。β-カテニン非依存性経路のなかで、とくに平面内細胞極性(planar cell polarity :PCP)と呼ばれるシグナル経路の構成因子であるPrickleはこれまでにゼブラフィッシュやアフリカツメガエルにおいて発生過程の収斂伸長運動にともなう細胞運動に密接に関与することが報告されていますが、その詳細な分子メカニズムは明らかにされていませんでした。
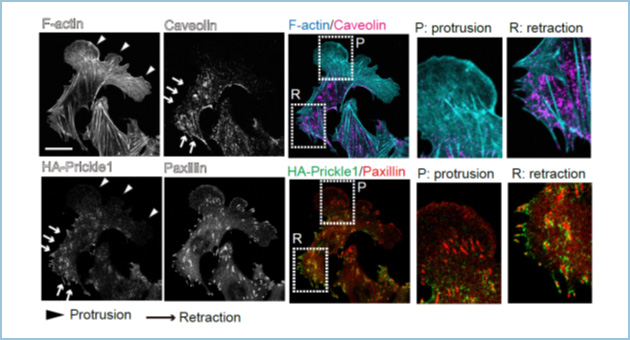
活発に細胞運動する胃癌細胞株において、Prickle1を発現抑制(ノックダウン)すると細胞の前後方向の極性化と細胞運動が抑制されました。そこで、Prickle1の細胞内での局在を検討したところ、Prickle1は細胞運動する際の伸長する細胞辺縁部(伸長端)ではなく、退縮する側の細胞辺縁部(収縮端)に偏った局在をすることが分かりました。収縮端には大きく成長した接着斑が多数存在し、細胞が移動するために活発に接着斑が分解(ターンオーバー)されています。そこでPrickle1とパキシリン(接着斑構成タンパク質)の局在を検討したところ、Prikcle1は収縮端の接着斑近傍に特異的に集積していることが分かりました(図1)。さらに、Prickle1の接着斑近傍への局在は、接着班が形成されると現れ、接着班が分解されると消失することから、接着斑依存的であることが明らかになりました。Prickl1をノックダウンすると接着斑のターンオーバーが抑制されることから、Prickle1は細胞運動において収縮端の接着斑近傍に局在して接着斑のターンオーバーを促進することが明らかになりました(図2)。
図1.Prickle1の収縮端における接着斑近傍への局在
Prickle1(緑)は細胞の伸長端(Protrusion)ではなく、Caveolin(赤紫)陽性の収縮端(Retraction)側に集積し、接着斑(赤)の近傍に局在した。
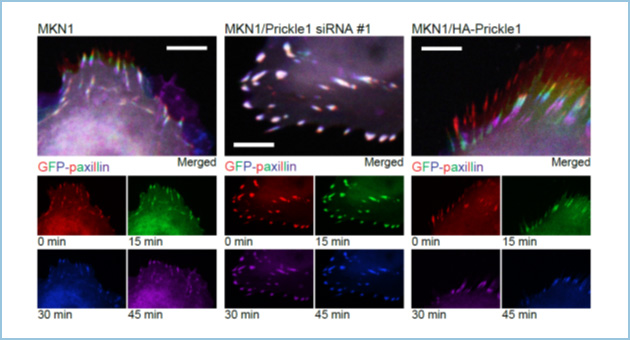
図2.Prickle1による接着斑のターンオーバーの制御
コントロール(左)、Prickle1ノックダウン(中央)、Prickle1過剰発現(右)細胞における接着斑(GFP-paxillin)を15分ごとに0分(赤)、15分(緑)、30分(青)、45分(紫)と観察し(下段小パネル)、重ねあわせた(上段大パネル)。Prickle1ノックダウンと過剰発現により接着斑のターンオーバーがそれぞれ抑制または促進した。
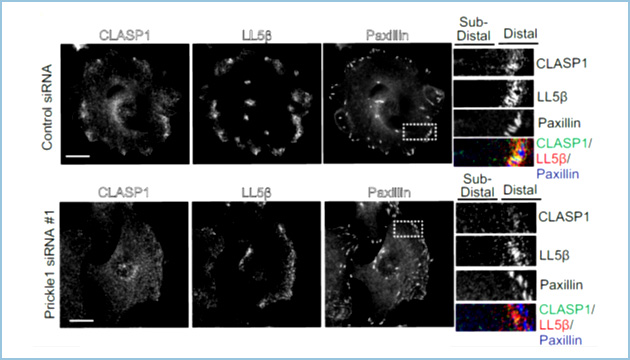
Prickle1と類似して、接着斑の近傍に特異的に局在することがこれまでに知られているタンパク質としてCLASP1/2(CLASPs)とLL5βがあります。CLASPsは微小管伸長端集積因子として知られており、LL5βとCLASPsの結合を介した微小管の接着斑へのリクルートが接着斑のターンオーバーを誘導することが明らかになっています。Prickle1はCLASP1およびLL5βと接着斑近傍で共局在し、互いに複合体を形成しました。Prickle1のノックダウンはLL5βの局在には影響しませんでしたが、CLASPの接着斑近傍への局在が損なわれ(図3)、さらに接着斑と微小管の相互作用が抑制されました。一方で、CLASPsのノックダウンはPrickle1の局在には影響しませんでしたが、LL5βをノックダウンするとPrickle1の接着斑近傍への局在が損なわれることが分かりました。すなわち、Prickle1はLL5β依存的に接着斑近傍に局在し、CLASPsの同部への集積と微小管の接着斑への相互作用に必要であることが明らかになりました。
図3.Prickle1のノックダウンが接着斑近傍へのCLASP1およびLL5βの局在に与える影響
Prickle1をノックダウンした細胞(下段)では接着斑(Paxillin, 青)近傍へのCLASP1(緑)の局在が損なわれた。一方、LL5β(赤)の接着斑への局在には影響しなかった。
Prickle1はC末端側にファルネシル基による脂質修飾を受けます。本脂質修飾はPrikle1が細胞膜および接着斑近傍へ局在すること、Prickle1による細胞運動・極性化および接着斑のターンオーバーの促進に必要でした。さらに、Prickle1とCLASP1、LL5βとの結合もPrickle1のファルネシル化に依存していました。一方、ファルネシル化の代わりに、N末端にミリスチン酸修飾配列を付加したPrickle1は、細胞膜へは局在するものの、接着斑近傍への集積とCLASP1およびLL5βとの結合が損なわれ、細胞運動・極性化と接着斑のターンオーバーを促進することが出来ませんでした。すなわち、Prickle1がCLASP1およびLL5βと複合体を形成し、接着斑近傍に局在して微小管を介して細胞運動を促進するためにはC末端側へのファルネシル化修飾が必要であることが明らかになりました。
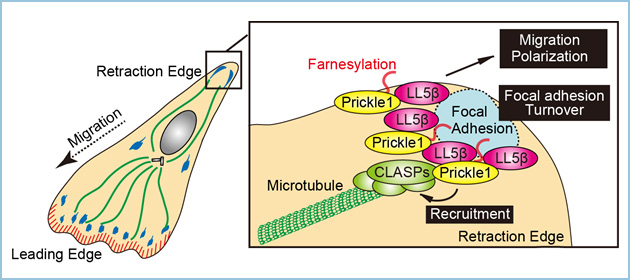
これらの結果から、細胞運動において、Prickle1がLL5βを介して収縮端の接着斑近傍に局在し、CLASPと微小管の接着斑への集積を促進することで接着斑のターンオーバーと細胞運動・極性化を促進する新たな機構が明らかになりました(図4)。
図4.モデル図
ファルネシル化修飾されたPrickle1がLL5βを介して収縮端の接着斑近傍に局在し、CLASPと微小管の接着斑への集積を促進することで接着斑のターンオーバーと細胞運動を促進する。
Wnt5aシグナルによる細胞運動制御
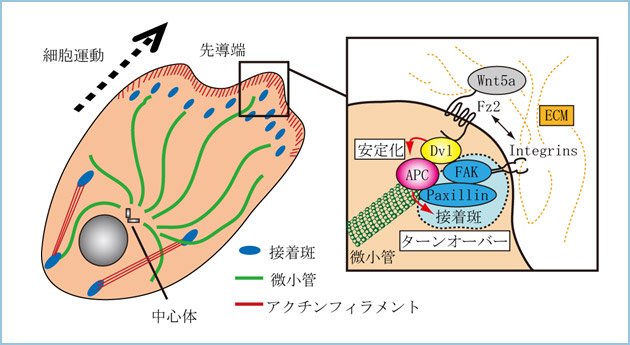
細胞が運動を始める際にまず起こるのは、アクチン重合による細胞の進行方向前方への伸展であり、細胞の前方に新しいフォーカルコンプレックスが形成されます。引き続き、すぐ後方の細胞接着斑が消失し、さらに細胞後方ではストレスファイバーの収縮が起こり、細胞が全体として前方に進行していきます。この一連の経過には低分子量G蛋白質RacやRho、Cdc42が重要な役割を果たします。アフリカツメガエル初期胚の原腸形成時の細胞運動においては、RacによるJNKの活性化が重要です。Wnt5aとそのシグナル経路(β-カテニン非依存性経路)がこの細胞運動制御に関与していることが明らかになりつつあります。私達は、Wnt5aはcPKCを介して細胞運動を促進し、細胞が運動する際に足場となる細胞接着斑の構成因子であるfocal adhesion kinase (FAK)やRacを活性化することを見出しました。また、Wnt5aの発現を抑制すると細胞運動先端部におけるラッフリングが抑制され、細胞接着斑のパキシリンのターンオーバーが阻害されました。さらに、細胞が運動するときに形成される先導端(リーディングエッジ)においてWnt5aの下流で、Dvlが微小管伸長端集積因子であるAPCと複合体を形成し、直接細胞表層での微小管の安定化を引き起こし、接着斑のターンオーバーを制御する可能性を示すデータを得ています(図1)。
図1.Dvl/APC複合体を介した細胞接着、細胞運動の制御
Wnt5aは先導端においてFz2受容体と結合し、Dvlと+TIPsとして知られているAPCとの複合体形成を促進する。私達は、Dvl/APC複合体が先導端において微小管を安定化させ、さらにFAKやPaxillinと相互作用し、細胞接着斑のターンオーバーを促進することにより細胞運動を制御することを示している。
Wnt5aシグナルによる細胞極性決定機構
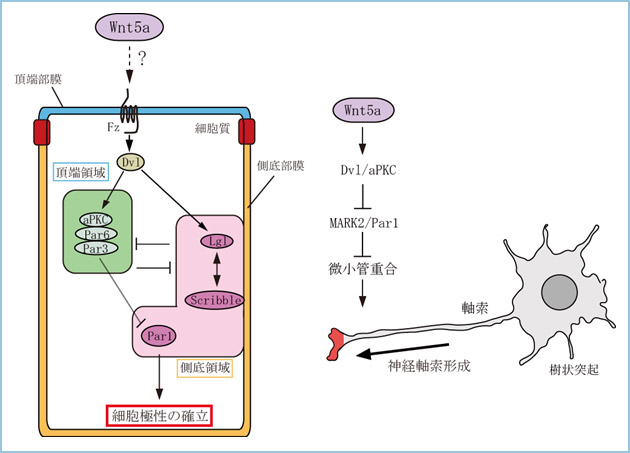
一般に、細胞極性とは細胞が形態的あるいは機能的に非対称性を示すことであり、分化した上皮細胞や神経細胞に特有の形態と機能は哺乳動物細胞における細胞極性の代表的な例です(図1)。上述した方向性を持った細胞運動も細胞が極性化した結果起こったと考えられます。上皮細胞には頂部-基部軸に沿った極性に加えて、頂部-基部軸と直交する平面に沿った平面内細胞極性 (Planar Cell Polarity, PCP) が存在します。WntとPCPの関連はよく知られていますが、上皮細胞においてWntシグナルが頂底極性を制御しているかは明らかにされていません。私達は、腎上皮細胞(MDCK)や胆管上皮細胞(HPPL)の三次元培養を用いてCyst(のう胞)形成時の頂底極性決定におけるWntシグナルの役割を解析を行っています。
図1.Wnt5aによる上皮細胞極性および神経軸索形成の制御
(左)上皮細胞の頂底極性形成機構。Dvlは頂端領域と側底領域の各々に特異的な局在を示す分子群の両方に作用する。最終的には、本来側底部に局在するPar1の活性が、頂端部では抑制されることによって上皮細胞の頂底極性が形成される。
(右)Wnt5aによる神経軸索形成の制御。Dvl/aPKCが軸索先端領域でのMARK2/Par1を阻害することによって軸索が形成される。
LRP6-Fz2-Dvl2-APCシグナル軸による細胞周期調節機構
Wntシグナル経路を構成するタンパク質は、細胞周期においてシグナル伝達因子として下流の遺伝子発現調節や細胞増殖制御に関与するだけでなく、細胞分裂期においても重要な機能を有することが明らかになりつつあります。私共は、Wntシグナル経路において中心的な役割を担うDvlホモログのひとつDvl2が有糸分裂期において多彩な機能を有することを見出しました。Dvlは、神経細胞において微小管の安定化に寄与することや線虫初期胚の不均等分裂時に紡錘体軸を制御することが報告されていましたが、哺乳類動物細胞の細胞分裂期における機能は明らかになっていませんでした。
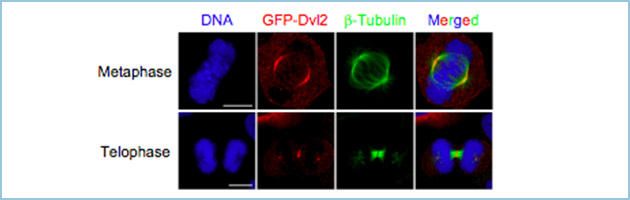
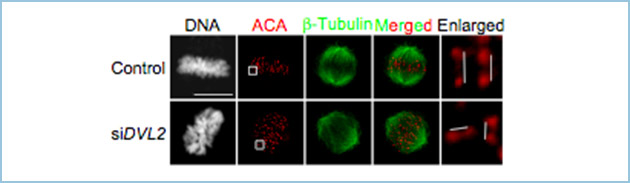
Dvl2は分裂前期から分裂中期までは紡錘体極、及び、その近傍の微小管に強く局在し、動原体への局在が認められました(図1)。また、微小管脱重合剤・ノコダゾールにより紡錘体チェックポイントの活性化を誘導すると、動原体への局在が顕著に観察されました。さらに、分裂終期では分裂溝に強く局在しました(図1)。
図1.有糸分裂期におけるDvl2の細胞内局在
GFP-Dvl2は分裂前期から分裂中期までは紡錘体極、及び、その近傍の微小管に強く局在し、動原体への局在も認められた。また、分裂終期では分裂溝に強く局在した。
Dvl2ノックダウン細胞を用いた表現型解析から、Dvl2は紡錘体軸、微小管-動原体結合、及び、紡錘体チェックポイントの制御に関与することが明らかになりました(図2、3、4)。有糸分裂期キナーゼのひとつであるPolo-like kinase 1(Plk1)によりDvl2はリン酸化され、そのリン酸化部位は206番目のスレオニン(T206)であることがわかりました。Dvl2の非リン酸化型変異体T206Aを用いて解析を行いますと、Plk1によるDvl2のリン酸化が紡錘体軸、及び、微小管-動原体結合の制御に必要でした。一方、Dvl2はリン酸化非依存的に紡錘体チェックポイントキナーゼであるMps1の活性化、及び、Bub1とBubR1の動原体集積に関与し、紡錘体チェックポイントを制御しました。
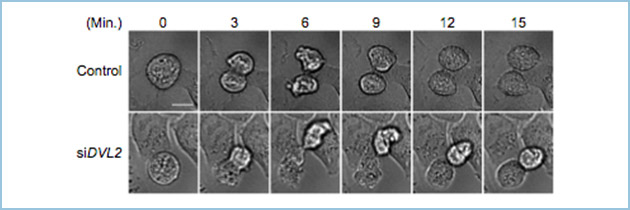
図2.Dvl2ノックダウン細胞で観察される紡錘体軸の異常
コントロール細胞では、分裂後期に移行した後に、プレート面に対して平行に細胞分裂を行う(写真上段)。一方で、Dvl2ノックダウン細胞では紡錘体軸の異常により、片方の娘細胞が上方に持ち上がる現象が観察される(写真下段)。
図3.Dvl2ノックダウン細胞で観察される微小管-動原体結合の異常
Dvl2ノックダウン細胞では、微小管-動原体結合が減弱し、微小管からの張力が減少するため、姉妹動原体間の距離が短くなり(写真・右端)、染色体の赤道面への整列が十分ではない(写真・左端)。
Dvl2ノックダウン細胞で観察される紡錘体軸、及び、微小管-動原体結合の異常はAPCノックダウン細胞においても観察されました。また、Dvl2は細胞周期を通じてAPCと複合体を形成することから、表現型の類似性と合わせて、Dvl2はAPCと複合体を形成し、紡錘体軸、及び、微小管-動原体結合を制御する可能性が考えられました(図4)。
さらに、紡錘体軸については、Wntの受容体であるFz2とβ-catenin 依存性経路の共役受容体であるLRP6が必要でしが、β-catenin/Tcf(Lef)の転写活性化能は必要ではありませんでした。LRP6-Fz2は有糸分裂期において細胞膜上に局在することから、LRP6-Fz2-Dvl2-APCからなる新しいシグナル経路が細胞表層と星状糸の結合を制御している可能性が考えられました(図4)。
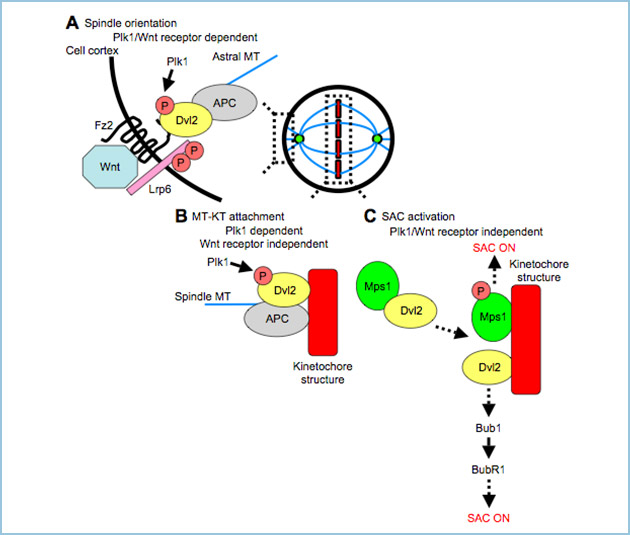
図4.モデル図
A. Dvl2による紡錘体軸の制御において、Plk1によるリン酸化、及び、Wnt受容体(Fz2、LRP6)の関与が示唆された。また、Dvl2はAPCと複合体を形成し、紡錘体軸を制御することが示唆された。
B. Dvl2による微小管-動原体結合の制御において、Plk1によるリン酸化の関与が示唆された。一方、Wnt受容体は微小管-動原体結合の制御には関与しなかった。
C. Dvl2は、Plk1によるリン酸化非依存的及びWnt受容体非依存的に紡錘体チェックポイントを活性化し、さらに、紡錘体チェックポイントキナーゼであるMps1の活性化、及び、Bub1とBubR1の動原体集積に関与することがわかった。
Wnt5aシグナルによる細胞質分裂制御機構
上述しました通り、Wntシグナル伝達による細胞分裂期前期から中期における役割は明らかになりつつあります。一方、私たちは分裂期後期におけるWntシグナルの機能にも注目して解析を行っています。
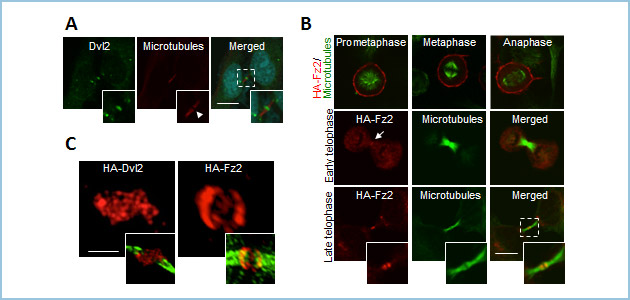
Dvlは細胞質分裂期においてMidbodyに局在することを私たちは見いだしました(図1A)。大変興味深いことにWntシグナルのリセプターであるFz2は細胞分裂期前期から終期にかけて細胞表層に存在しますが、細胞質分裂時にはmidbodyに局在しました(図1B)。超高解像度顕微鏡を用いた観察によって、midbodyにおいてDvl2とFz2はそれぞれweb様構造とリング状構造を形成することがわかりました(図1C)。
図1.Dvl2とFz2の細胞質分裂期におけるmidbodyへの局在
A. Dlv2(緑色)は細胞質分裂期にmidbody(微小管、赤色)の中央に局在する。青色;DNA。
B. Fz2(赤色)は細胞分裂期前期から終期にかけて細胞膜上に局在するが、分裂期後期から細胞質分裂期においてmidbodyの中央に局在する。緑色;微小管。
C. 超高解像度イメージによる解析によって、赤色で示しているDvl2とFz2はそれぞれweb様の構造とリング状の構造をとることが分かる。
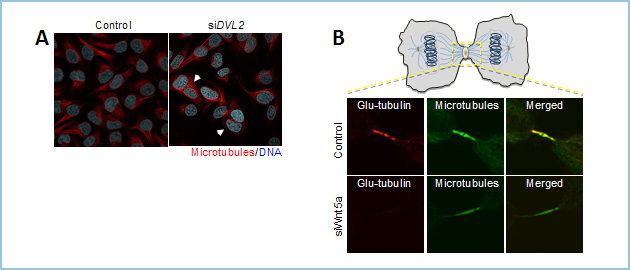
細胞質分裂が障害されると多核になることが知られています。そこで、siRNAを用いたノックダウン実験を行ったところ、Dvl及びWntβ-カテニン非依存性経路の代表的リガンドとリセプターであるWnt5a、Ror2、Fz2の発現抑制により多核細胞の増加を認めました(図2A)。
細胞質分裂時に形成されるMidbodyは高度に安定化した微小管から構成され、細胞質分裂に必要な構成因子の集積に必要であることが知られています。Wnt5aシグナルは微小管の安定性に関与することが示唆されていることから、そのノックダウンによる微小管への影響を解析したところ、midbody微小管の安定性が減弱していることがわかりました(図2B)。さらに、midbodyへリクルートされてくるタンパク質群を解析した結果、midbodyの脱離(abscission)に関与することが知られているESCRT-IIIタンパク質の局在が損なわれることを見いだしました。さらにESCRT-IIIはFz2とmidbodyにおいて共局在し、Wnt5a依存的にFz2と結合することがわかりました。また、Fz2のmidbodyへの局在はWnt5aやDvl、さらにRab11のノックダウンによって損なわれることから、微小管を基盤とした細胞内タンパク質輸送によって制御されていることが考えられました
図2.Wnt5aシグナルの異常に基づく細胞分裂期の表現型
A. HeLaS3細胞においてDvl2をノックダウンすると多核細胞(A右の写真、矢頭で示す細胞)が増える。
B. Wnt5aシグナルのノックダウンによりmidbody微小管の安定性が低下する。
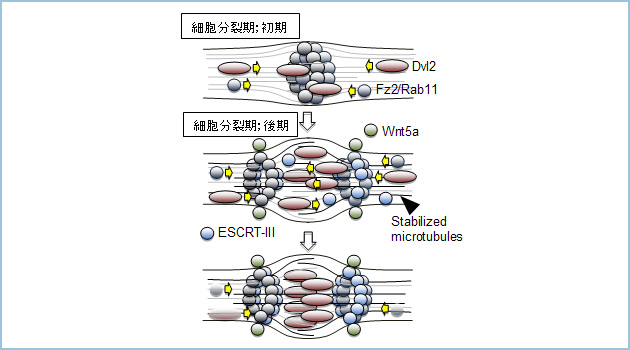
これらのことから、細胞質分裂期におけるWnt5aシグナルの役割について次のようなモデルを考えています(図3、モデル図参照)。
分裂期終期に、Fz2はRab11によってmidbodyへリクルートされてきます。その後、Wnt5aおよびそのリセプターであるFz2、Ror2からのシグナルはDvlをmidbodyへリクルートし微小管を安定化します。微小管の安定化はRab11によるFz2の輸送をサポートすると考えられます。さらにWnt5a刺激はmidbodyにおけるFz2とESCRT-IIIとの結合を誘導することによって、ESCRT-IIIの局在を制御し、ESCRT-IIIによる脱離をサポートするのではないかと考えています。
図3.モデル図