Wntと増殖因子シグナルによる上皮形態形成
線維芽細胞MARK1による肺胞上皮組織の形態形成制御
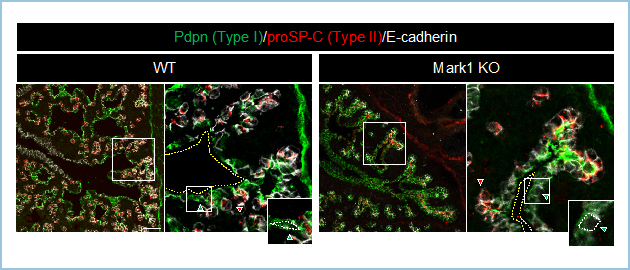
気管上皮が分岐構造を形成後、形成後期(E16.5以降)に気管支遠位側が肺胞上皮細胞分化を伴いながら肺胞を形成し、肺の基本構造を獲得します。この過程にも、Fgf、Wntなど増殖因子シグナルを介する上皮-間葉の相互作用が分岐および肺胞形成に重要であることが明らかになっています。しかし、増殖因子シグナルがどのように上皮組織の動態を制御し、肺胞を形作るのか不明でした。その分子機構を解析する過程で、上述のMark1は発生後期になると発現が線維芽細胞特的に限局しました。さらに、トリプルCRISPR法を用いて作製したMark1 ノックアウト(KO)胎児肺では気管支遠位側に形成される肺胞内腔の拡張が阻害されました(図1)。
図1.E17.5 Mark1 KO肺。Pdpn: I型肺胞上皮マーカー、proSP-C: II型肺胞上皮マーカー。Mark1 KO 胎児肺では気管支遠位側のI型肺胞上皮の進展と肺胞内腔の拡張が阻害された。
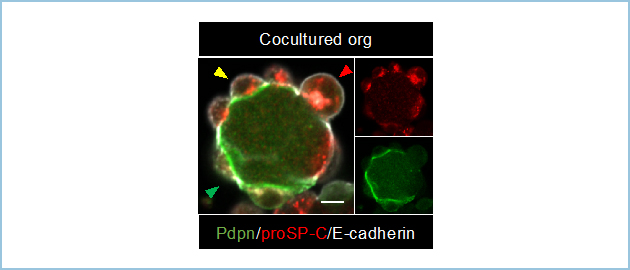
そこでMark1 の機能解析を通じて、発生後期の肺間葉、特に線維芽細胞が肺胞形成をどのように制御するのかを明らかにすることを試みました。まず胎児期における肺胞内腔の拡張機構を解析するため、胎児肺由来上皮細胞を用いた肺胞オルガノイド と線維芽細胞の三次元共培養系を構築しました。本培養系で作製したオルガノイドでは、胎児期同様二種類の肺胞上皮細胞と肺胞前駆細胞が認められました(図2)。
図2.胎児肺由来線維芽細胞と共培養した胎児肺由来肺胞オルガノイド。
肺胞オルガノイドと線維芽細胞の三次元共培養により、肺胞上皮細胞と肺胞前駆細胞が出現した。
胎児Mark1 KO肺由来線維芽細胞との共培養では肺胞オルガノイドにおける肺胞上皮細胞の分化には影響はありませんでしたが、オルガノイド内腔の拡張が抑制されました。詳細な解析から、Mark1は線維芽細胞のシリアを制御することでHedgehogシグナルを適正化し、線維芽細胞の活性化を誘導することが判明しました。その結果、肺胞上皮細胞の基底側コラーゲンが蓄積することで肺胞上皮細胞の扁平化を誘導し、内腔が拡張する可能性が示唆されました。そこで、数理モデルを用いて内腔拡張における細胞の扁平化の役割を解析したところ、細胞の扁平化は内腔拡張の必要条件でしたが、パラメータとしては不十分でした。そこで既報をもとづき本モデルに内圧を加えたところ肺胞拡張が再現されました。
以上のことから、線維芽細胞はコラーゲン蓄積を介して間質をリモデリングすることで肺胞上皮細胞を扁平化し、羊水等による内圧に依存して肺胞拡張を促進する作用があることが示唆されました。
気管支から肺胞が形成される仕組み
肺、膵臓、唾液腺などの管腔臓器は、ガス交換や分泌物の産生など臓器固有の機能を有する「腺房」とその通り道である「導管部」により構成されます。肺では導管部である気道が発生前期−中期(分岐形成期)に分岐し、腺房部である肺胞が発生後期から生後(肺胞形成期)に形成され成熟型臓器となります。気道の形態形成は規則的なパターンに従って分岐することが知られており、他の管腔臓器に通じる形態形成の原理を追求するモデルとして、これまで多くの生物学者や数理学者の探究心を刺激し、様々なことが明らかになってきました。しかし、発生後期になり気道遠位端に肺胞が誘導されるメカニズムは未解決な問題でした。この問題に対し、「シグナルネットワーク」と「細胞形態変化」に注目し、解析を試みました。
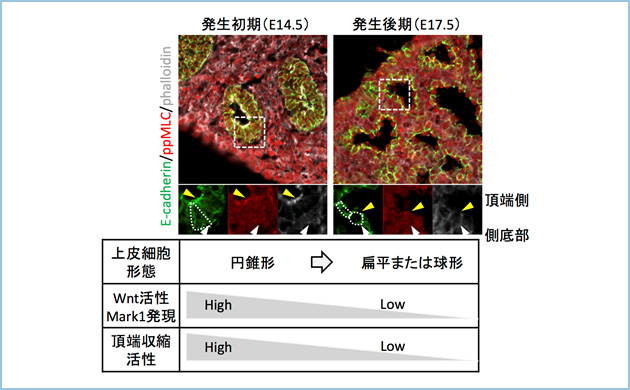
まず、分岐形成期での上皮細胞は頂端収縮し円錐形であるのに対し、肺胞形成期では頂端収縮が起こらず細胞の形状も立方形(二型肺胞上皮細胞)あるいは扁平(一型肺胞上皮細胞)でした。上皮組織に特異的に発現するWntリガンドや受容体及び標的遺伝子などWntシグナル構成因子の発現を解析したところ、分岐形態形成期において発現が高く、肺胞形成期になると発現が低下していました。安定型β-cateninをE17.5マウスに発現させ、肺胞形成期にてWntシグナルをin vivoで恒常的に活性化すると、頂端収縮が誘導され、肺胞形成が阻害されんました。すなわち、Wntシグナルの抑制が肺胞形成に必要であることがわかりました(図1)。
図1.Wntシグナルによる分岐・肺胞形成の制御機構
肺分岐形成期においてはWntシグナルによって頂端収縮に伴い分岐が誘導された。それに対し、肺胞形成時期にはWntシグナル活性が低下し、頂端収縮活性の低下と共に肺胞が形成された。
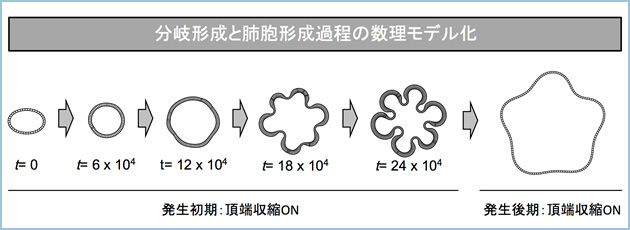
次に、細胞形態と組織形状との関連を数理モデルにて解析したところ、一層の細胞塊(シスト)で個々の細胞を頂端収縮させるとシストが自律的に分岐するのに対し、頂端収縮しない場合は分岐形態を形成できませんでした。さらに、同モデルにおいて、全周性に基底側方向への運動を誘導したところ、頂端収縮しないシストでは肺胞様構造が形成されたが、頂端収縮する場合は肺胞様構造の形成が阻害されました(図2)。
図2.分岐・肺胞形成過程の数理モデル化
肺上皮組織を一層のシストと見なし、頂端側に収縮を誘導すると分岐が形成され、その後収縮を除去すると、肺胞様構造が誘導された。
すなわち(1)分岐形成期ではWntシグナルによって上皮細胞が頂端収縮すると分岐構造が形成されること、(2)肺胞形成期ではWntシグナル低下に伴って頂端収縮活性が低下し、細胞が立方形あるいは扁平化することにより肺胞構造へと変化すると考えられました。さらに、分岐形態形成期の上皮組織におけるWntシグナルの下流因子としてMAP/microtubule affinity-regulating kinase 1(Mark1)を同定し、肺上皮組織培養系を用いてMark1の発現抑制実験によって、Wntシグナルの下流でMark1が頂端収縮を制御することを見出しました。
WntとKITシグナルの協調による唾液腺の形づくりと機能獲得過程の制御
発生過程において、肺や腎臓、腸管、唾液腺等の私たちの体を構成している多くの臓器は、上皮細胞が集団として活発に増殖しながら立体的な管状の構造(上皮管腔構造)を形成し、その後臓器固有の細胞へと分化することによって機能を獲得します。私たちはこれまでに培養上皮細胞や腎臓などの管腔臓器原基の多次元的な培養法を駆使して、液性因子シグナルにより制御される上皮管腔構造の形成機構(“形づくり”)を明らかにしてきました。しかし、器官形成の前期にみられる“形づくり”だけでなく、後期にみられる“分化(機能獲得)”へと経時的に進行していく一連の発生過程を適切に調節する機構についてはいまだ明らかにされていません。そこで本研究では、外分泌腺の一種である唾液腺の発生をモデルとして“形づくり”から“分化”へと移行する過程を制御するシグナル機構について検討しました。
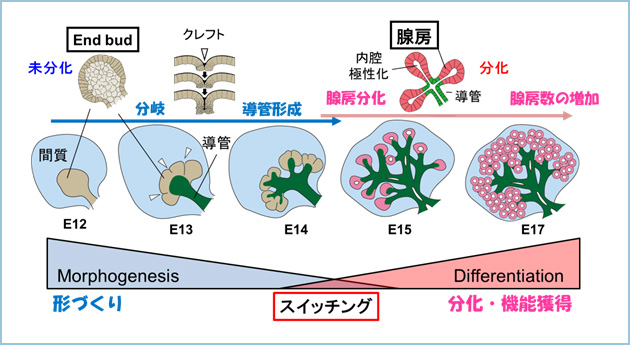
唾液腺は発生過程において、未分化な終末部上皮(end bud)が活発に分岐をしながら管腔(導管)構造を形成し、その後唾液を産生する腺房へと急速に分化します(図1)。
図1.マウス唾液腺の発生過程における“形づくり”と“分化”
唾液腺の発生過程においては、分岐をともなって管腔構造を形成する“形づくり”とend budが腺房構造へと変化する“分化・機能獲得”が経時的に進行する。
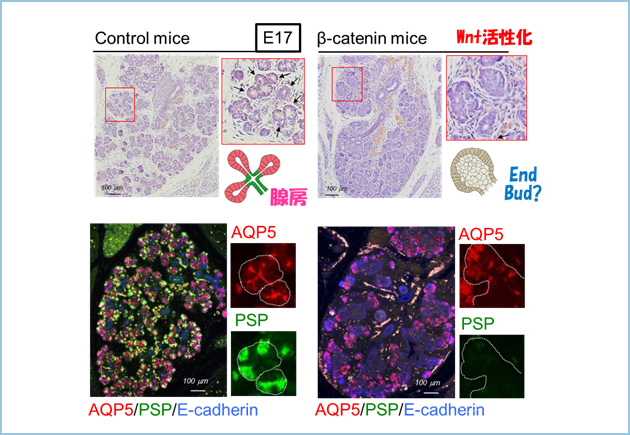
私たちは発生過程において重要な液性因子シグナルのひとつであるWntシグナルを恒常的に活性化したマウス(安定型β-カテニン発現マウス、Ctnnb1lox(ex3))の唾液腺において、器官の大きさに異常は認めないものの、胎生17日目における腺房分化が著しく抑制されていることを見出しました(図2)。
図2.Wntシグナルを活性化したマウス(胎生17日目)における腺房分化不全
Wntシグナルを恒常的に活性化した胎生17日目のマウス(安定型β-カテニン発現マウス、Ctnnb1lox(ex3))の唾液腺の腺房分化マーカーAQP5(赤)と腺房分泌タンパク質PSP(緑)を染色した。コントロールの唾液腺ではAQP5(赤)とPSP(緑)が強く発現していたが、Wntシグナルを活性化した唾液腺ではその発現が抑制された。
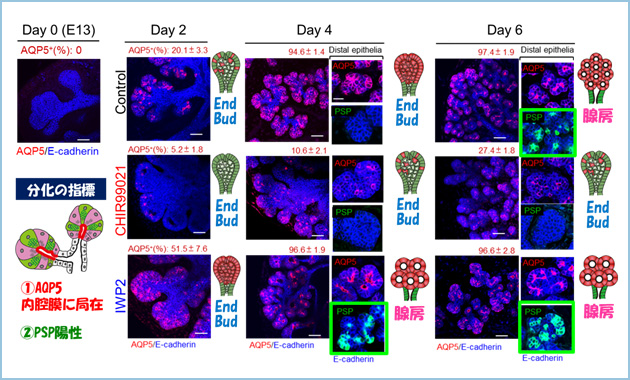
胎生13日目の唾液腺原基を摘出して器官培養すると、非極性化上皮からなるend budは培養4日目から6日目にかけて急速に極性化し、多房性の腺房構造を形成するとともに腺房分化マーカーの発現が強く上昇しました。一方、Wntシグナルの標的遺伝子であるAxin2の発現は腺房分化にともなって経時的に減少していました。器官培養において、Wntシグナルを恒常的に活性化させると、培養6日における腺房構造の形成が強く抑制され、end budは未分化な非極性化状態に維持されるとともに、導管の形成が促進しました。一方、Wntシグナルを阻害すると、end budはコントロールと比較して早期に極性化して腺房構造を形成するとともに、導管の形成が抑制されました(図3)。
図3.Wntシグナルによる腺房分化のタイミングの制御
胎生13日目のマウス唾液腺を摘出し、CHIR99021(Wntシグナル活性化)およびIWP2(Wntシグナル阻害)の存在下で器官培養した。CHIR99021処理によって、腺房分化が抑制され、end budが維持されたのに対して、IWP2処理は早期の腺房分化を誘導した。
さらに唾液腺発生の前期において、Wntシグナルの活性化は導管部ではなく、end budにおける細胞増殖を促進し、Sox10+またはKRT14+の前駆細胞数を増加させました。そこで、end bud細胞を特異的に蛍光標識して追跡したところ、end bud由来の細胞が導管を形成する様子が観察され、さらにWntシグナルの活性化はend bud由来の導管形成を強く促進することが明らかになりました。
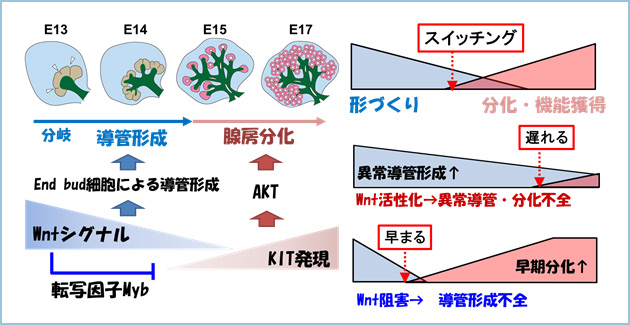
Wntシグナルによる腺房分化の制御メカニズムを検討したところ、発生後期ではWntシグナル活性の減弱にともなって、end budにおいてSCF受容体であるKITの発現が局所的に上昇し、AKTの活性化を介して腺房分化が誘導されていました。一方、end budに由来する導管形成は発生後期では認められませんでした。したがいまして、Wntシグナルは発生前期において、KITの遺伝子発現を抑制することによってAKTの活性を抑制し、腺房分化の進行を抑えていることが明らかになりました(図4)。本研究から、WntとKITシグナルの活性化バランスが器官形成過程における“形づくり”から“分化(機能獲得)”へのスイッチングを調節する新たな機構が明らかになっています。
図4.WntとKITシグナルの活性化バランスが“形づくり”と“分化”のスイッチングを制御する
Arl4cの発現を介する上皮管腔組織形成
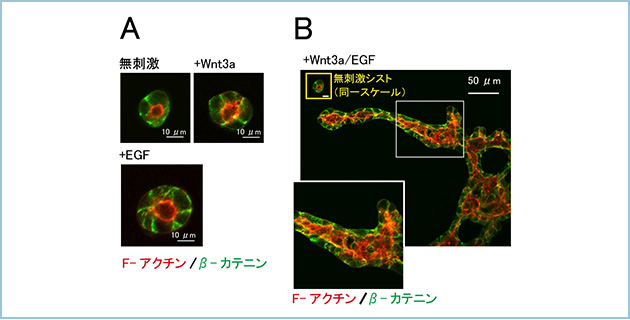
腎臓や肺をはじめとする管腔臓器の発生には、β-カテニン経路も重要な役割を果たすことが知られています。in vitroでの新たな管腔形成モデルを確立するためにラット腸管上皮細胞(IEC6)を基底膜タンパク質(マトリゲル)内で三次元的に培養しました。ある種の上皮細胞はECMゲル内で三次元培養すると、基質との接着面が基底側、その反対側は頂端側となり極性化して、内腔を有した球状の構造、すなわちcyst(シスト)を形成します。IEC6細胞はマトリゲル内で増殖や運動を停止し、小さなシストを形成しました。EGFやHGF、Wnt3a、Wnt5a等の液性因子(増殖因子)は単独では、IEC6細胞のcystの形状にほとんど影響しませんでした(図1A)。そこでこれらの増殖因子を組み合わせ作用させてみたところ、EGFとWnt3aによりIEC6細胞のcystの形状は著しく変化し、チューブ状の上皮管腔構造が形成されました(図1B)。
図1.三次元マトリゲル基質中でのWnt3aとEGFシグナルによる上皮管腔構造形成
A. IEC6細胞は三次元マトリゲル中で頂底極性化して内腔を有する小さなシストを形成する。Wnt3aやEGFのような液性因子(増殖因子)はマトリゲル中のシストの形態に大きな影響を与えなかった。
B. Wnt3aとEGFで同時に刺激を行うとシスト構造は劇的に変化し、伸長化してチューブ状の管腔構造を形成した。
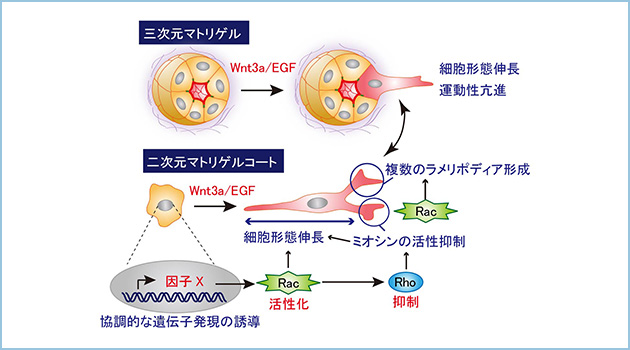
EGFとWnt3aは相乗的に作用して、ある遺伝子の発現を誘導しました。この遺伝子産物は、細胞骨格を制御する低分子量タンパク質Racの活性化を引き起こし、更にはRhoの活性を抑制しました。IEC6細胞はRacとRhoの活性が調節されることにより細胞骨格が変化し、三次元基底膜基質中で形態を伸長変化し、運動能が亢進しました(図2)。
図2.Wnt3aとEGFシグナルの協調による上皮細胞の細胞骨格・形態制御と運動能の活性化
三次元マトリゲル中のIEC6細胞に対してWnt3aとEGFによる刺激を行うと細胞が基質側へ向けて伸長化して運動(移動)能が亢進する。マトリゲルコートしたディッシュ(二次元マトリゲルコート)上でも上皮細胞はWnt3aとEGFの刺激によって伸長化し、複数のラメリポディアを形成した。また先導端部周囲のミオシンの活性が抑制されていた。Wnt3aとEGFは遺伝子Xの発現を協調的に誘導し、局所的なRacとRhoの活性を調節することで細胞骨格と形態を制御し、三次元基底膜基質中での上皮細胞の運動能を活性化した。
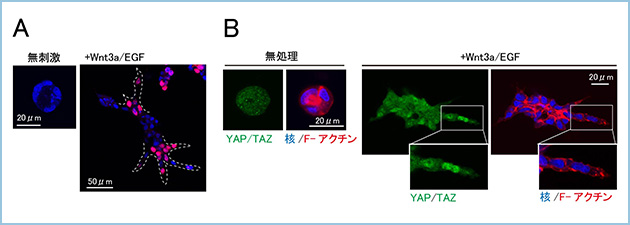
シストを構成するIEC6細胞は非常にコンパクトな形態を有していて増殖能が強く抑制されていましたが、Wnt3aとEGFによって管腔構造を形成する時は細胞増殖が活性化しました。この細胞増殖の活性化はWnt3aとEGFによって細胞形態が変化し、運動能が亢進している伸長部領域の細胞において強く認められました(図3A)。三次元基質中での細胞骨格や細胞形態の変化を細胞増殖のシグナルとしてとらえる役割を転写活性化因子であるYAPおよびTAZ(YAP/TAZ)が担っていました。YAP/TAZは細胞の形状や密度を感知して、細胞増殖を制御します。三次元基質中のIEC6細胞においてYAP/TAZは細胞質に局在し、細胞増殖は停止しましたが、Wnt3aとEGFによって細胞骨格と形態が変化することによりYAP/TAZが核内へ移行し、増殖シグナルを活性化する結果、管腔構造を形成すると考えられました(図3B)。
図3.YAP/TAZを介した管腔構造形成にともなう細胞増殖制御
A. IEC6細胞は三次元マトリゲル中のシストにおいてEdu(赤)をほとんどと取り込まず、細胞増殖は停止していた。一方で、Wnt3aとEGFによって管腔構造の形成が誘導されると、伸長部領域の細胞においてEduの取り込みが認められ、細胞増殖が活性化していた。
B. 三次元マトリゲル中でシストを形成するIEC6細胞はコンパクトな形態をとり、YAP/TAZは細胞質に局在した。一方で、Wnt3aとEGFによって管腔構造の形成が誘導されると、伸長部領域の細胞(形態が変化している細胞)においてYAP/TAZの核内移行が認められた。
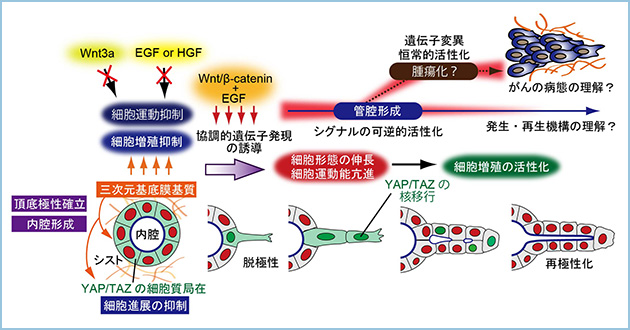
これらの結果から、液性因子シグナルの協調的作用により、生体内では抑制されている上皮細胞の増殖や運動が解除され、細胞形態の変化が細胞周期を進行させることにより、上皮管腔組織を形成する分子機構の存在が示唆されました(図4)。
図4.モデル図
P2Y2受容体の発現を介する上皮管腔組織形成
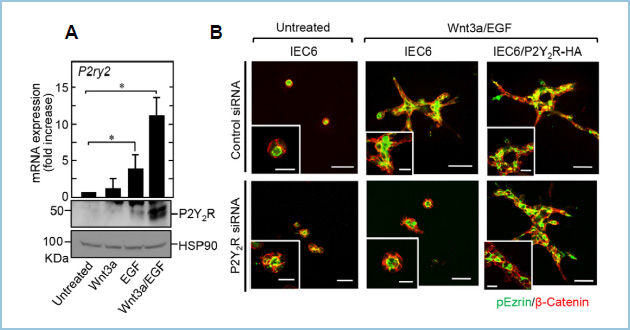
私共はこれまでに、正常ラット腸管上皮細胞(IEC6細胞)が三次元基質培養環境下でWnt3aとEGF(Wnt3a/EGF)の同時刺激によって上皮管腔構造を形成することを見出しています。Wnt3a/EGFの刺激によって協調的に発現が誘導され、管腔形成を制御する遺伝子としてこれまでにArl4cを報告していますが、今回新たに細胞外ヌクレオチド(ATP)受容体であるP2Y2 receptor(P2Y2R)を同定しました(図1A)。
P2Y2Rは細胞膜上の7回膜貫通型受容体で、細胞外ヌクレオチドであるATPまたはUTPをリガンドとします。P2Y2Rはこれまでに、血管内皮細胞や神経細胞などにおける機能が知られていますが、上皮管腔形成における役割については明らかになっていません。そこで、P2Y2Rの上皮管腔形成における役割についてIEC6細胞の管腔形成をモデルにして解析しました。
IEC6細胞においてsiRNAを用いてP2Y2Rを発現抑制するとWnt3a/EGF依存的な管腔形成が抑制されました。また野生型P2Y2Rの安定発現細胞株(IEC6/P2Y2R-HA)を樹立したところ、発現抑制の表現型が回復しました。さらに、IEC6/P2Y2R-HA細胞においてEGF単独存在下で管腔形成が誘導されました。これらの結果から、IEC6細胞における上皮管腔形成にP2Y2Rが必要であることが明らかとなりました(図1B)。
図1.IEC6細胞におけるWnt3a/EGF依存性管腔形成の新規制御因子P2Y2Rの同定
A. Wnt3aおよびEGFの共刺激によってP2Y2Rの発現が誘導される。
B. P2Y2RはWnt3a/EGF依存性管腔形成に必要である。
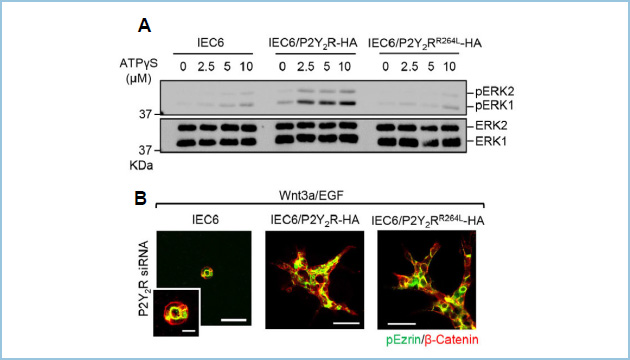
上皮管腔形成においてP2Y2Rのリガンドの関与を明らかにするために、リガンド不応性変異体(P2Y2RR264L)を作製しました(図2A)。予想に反して、IEC6/P2Y2RR264L細胞はIEC6/P2Y2R-HA細胞と同様に、P2Y2Rの発現抑制の表現型を回復しました。この結果から、IEC6細胞における上皮管腔形成にはP2Y2Rのリガンド応答性は必要ではないことが明らかになりました(図2B)。
図2.P2Y2Rリガンドの管腔形成への関与
A. リガンド不応性変異体P2Y2R(P2Y2RR264L)を作製し、IEC6細胞に安定発現させたところ、野生型P2Y2Rと比較してATPに対するERK活性化能が消失した。
B. P2Y2RR264Lの発現は野生型P2Y2Rと同様に、P2Y2Rノックダウンの表現型を回復した。
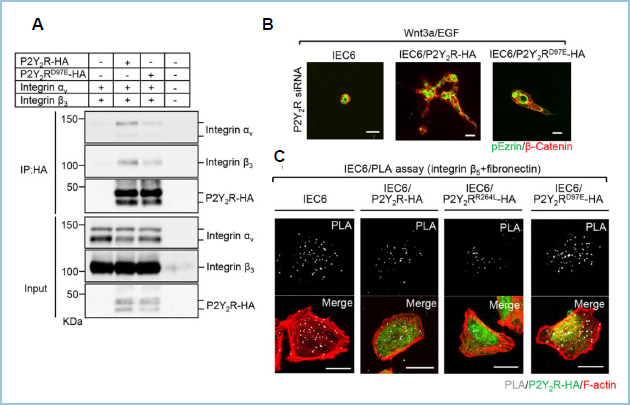
そこで、P2Y2Rのリガンド非依存的な機能が上皮管腔形成を制御している可能性を考え、P2Y2Rにユニークな特徴として細胞外領域にRGD(ラットではQGD)配列を有している点に注目をしました。既知の報告で、このRGD配列はインテグリンと相互作用することが知られており、P2Y2Rとインテグリンとの相互作用が上皮管腔形成に関与している可能性を検討しました。Proximity ligation assay (PLA)および免疫沈降法を用いた生化学的解析によってIEC6細胞においてP2Y2Rとインテグリンが相互作用することを確認後、過去の報告にならってインテグリン非結合変異体(P2Y2RD97E)を作製しました(図3A)。その結果、IEC6/P2Y2RD97E細胞は、P2Y2Rの発現抑制の表現型を回復できませんでした(図3B)。さらに、P2Y2RD97EにはEGF存在下での管腔形成誘導活性がありませんでした。これらの結果から、IEC6細胞において上皮管腔形成の誘導にはP2Y2Rとインテグリンとの相互作用が必要であることが明らかになりました。
図3.P2Y2Rとインテグリンの相互作用の管腔形成への関与
A. P2Y2Rはインテグリンαvおよびインテグリンβ3と相互作用し、P2Y2RD97E変異体ではインテグリンとの相互作用が抑制された。
B. インテグリン非結合変異体P2Y2R(P2Y2RD97E)の発現はP2Y2Rノックダウンの表現型を回復出来なかった。
C. 野生型P2Y2Rとリガンド不応性変異体P2Y2Rはインテグリンとフィブロネクチンとの相互作用を抑制した。一方で、インテグリン非結合変異体P2Y2Rはインテグリンとフィブロネクチンとの相互作用に影響しなかった。
次に、P2Y2R とインテグリンの相互作用が上皮管腔形成を誘導するメカニズムについて検討しました。RGD配列は本来、インテグリンのリガンドである細胞外基質(ECM)タンパク質中に認められることから、P2Y2Rの発現がECMとインテグリンとの結合を競合的に阻害する結果、管腔形成が誘導される可能性を考えました。実際に、RGDペプチドを用いたインテグリン依存性細胞接着の抑制は、P2Y2Rの発現と同様に管腔形成を誘導しました。IEC6細胞におけるRGD型のECMタンパク質の発現を検討したところ、フィブロネクチンが高発現していました。そこで、フィブロネクチンを発現抑制(タンパクレベルで約50%程度)したところ、P2Y2Rの発現と同様の管腔形成が誘導されました。また、PLAおよび免疫沈降法を用いた生化学的解析によりP2Y2Rの発現がインテグリンとフィブロネクチンとの相互作用を抑制することを確認しました。さらに、IEC6細胞においてP2Y2Rの発現、またはRGDペプチドを用いたインテグリン依存性細胞接着の阻害は細胞の伸長形態変化を誘導しました。伸長形態変化した細胞において、細胞増殖活性化因子であるYAP/TAZが細胞質から核内へと移行している様子が観察されたことから、P2Y2RがYAP/TAZを介して管腔形成にともなう細胞増殖を活性化する可能性が示唆されました。
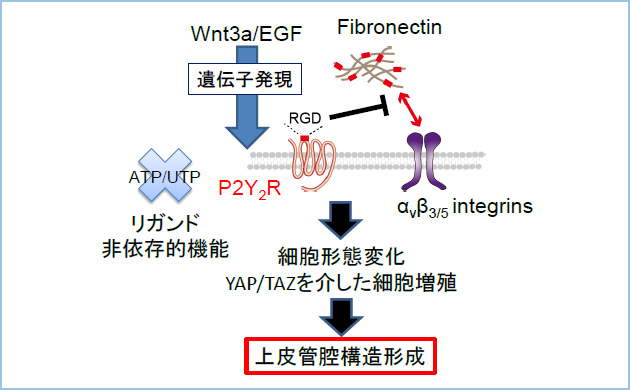
本研究によって、P2Y2Rはフィブロネクチンとインテグリンとの結合を適切に抑制することで管腔形成を誘導する新たな機構が明らかになりました。本研究成果は、種々の管腔臓器における発生と再生の仕組みの解明につながることが期待されます。また、WntとEGFシグナルの異常活性化にもとづく上皮管腔構造の破綻は種々のヒト癌において認められることから、P2Y2Rの発現が発癌と関連する可能性も示唆されました。
図4.P2Y2Rを介したWnt3a/EGF依存的上皮管腔形成の制御機構
Wnt3aとEGFシグナルにより協調的に発現誘導されるP2Y2Rはintegrinとfibronectinとの相互作用を抑制し、細胞の形態変化とYAP/TAZの核移行を介した細胞増殖を活性化し、上皮管腔形成を誘導する。
単一上皮細胞におけるWnt5aシグナルによる頂底極性制御
私共はラットの正常腸管上皮由来の細胞株であるintestinal epithelial cell 6 (IEC6細胞)を用いて、上皮細胞は細胞間接着がなくても単一細胞レベルで頂底極性を形成し、さらにこの極性化をWnt5aシグナルが促進することを明らかにしました。
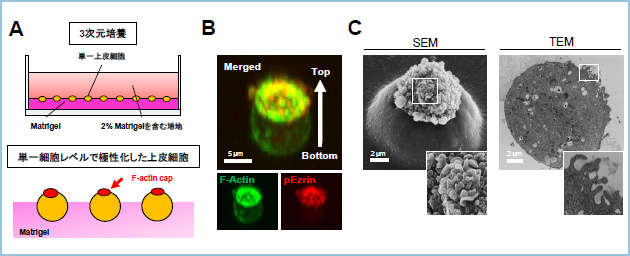
上皮細胞は頂底極性を形成しており、その極性の形成・維持には細胞外マトリックス(ECM)からのシグナルが重要です。Wntシグナルにおいてβカテニン非依存性経路は上皮細胞の平面内細胞極性を制御することが報告されていますが、頂底極性に対する役割は不明でした。そこで、私達はECMシグナルによる頂底極性形成制御に対するWntシグナルの役割を解析するために、IEC6細胞を用いて、3次元培養条件下に単一細胞レベルで頂底極性を形成する実験系を確立しました(図1)。頂底極性は細胞-ECM及び細胞-細胞間接着の二つによって形成されますが、この培養系を用いることで、細胞-ECM接着による上皮細胞極性化機構を解析することが可能となります。
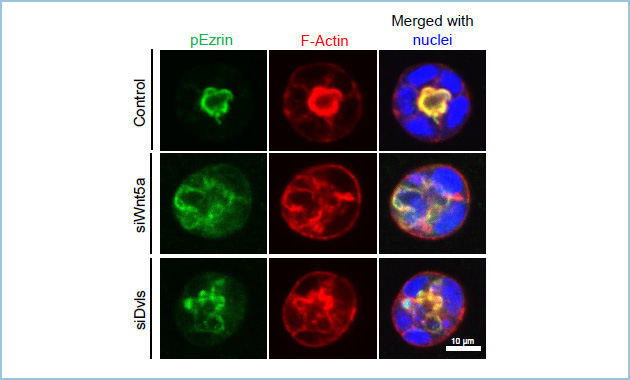
図1.3次元培養条件下で見られる単一細胞レベルでのIEC6細胞の頂底極性形成
A. 3次元培養のモデル図。
B. 単一細胞レベルで極性化したIEC6細胞において頂端部のマーカーであるリン酸化Ezrin(pEzrin, red)がF-actin(green)集積部において共局在する。
C.極性化した細胞を走査型電子顕微鏡(SEM)及び透過型電子顕微鏡(TEM)で観察したところ、F-actin capの部分には微絨毛様の構造が認められた。
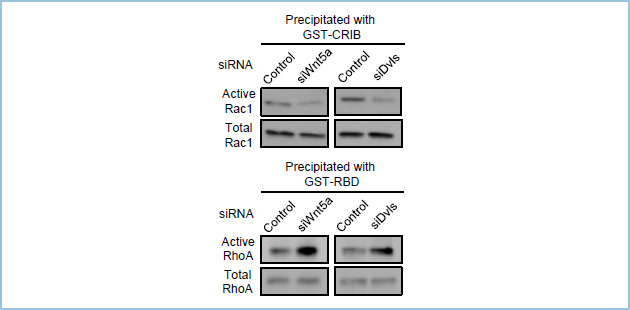
本実験系を用いた解析の結果、私達はECMにより誘導される単一細胞レベルでの上皮細胞の極性化にWnt5aシグナルが関与することを見出しました。さらに、Wnt5aシグナルはその下流で低分子量Gタンパク質Rac1の活性化を促進すると同時に、RhoAの活性化を抑制していることが明らかになりました(図2)。
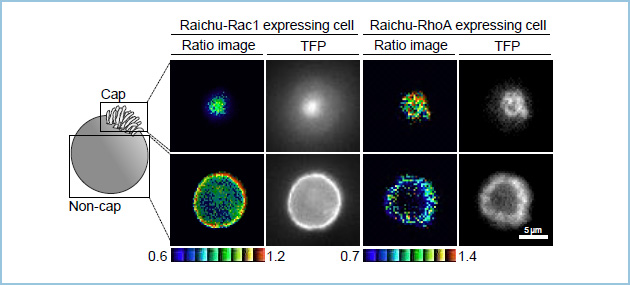
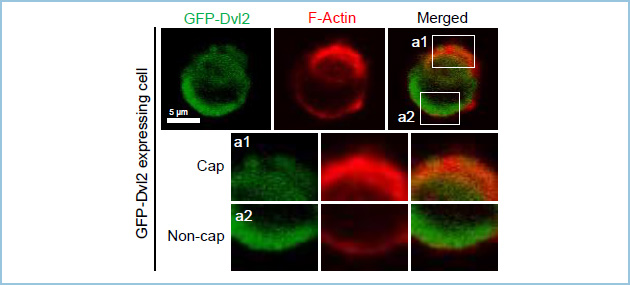
極性化した上皮細胞は非常に特徴的な非対称性の構造を取ります。そこで私達はRac1とRhoAそれぞれの活性の空間的な制御が極性化に重要と考え、その活性分布をfluorescence resonance energy transfer(FRET)を用いて解析しました。その結果、Rac1の活性は頂端部よりもむしろ基底部に相当する部分に強く見られ、RhoAの活性は頂端部により強く認められました(図3)。さらにWntシグナル構成因子であるDvl2が基底部に相当する部分に局在し、Tiam1(RacGEF)及びp190RhoGAP-A(RhoGAP)と複合体を形成しました(図4、5)。以上より、上皮細胞においてWntシグナルは低分子量G蛋白質制御因子を介してRac1とRhoAの活性を空間的に制御して、ECM接着誘導性の頂底極性形成に関与していると考えられました。
図2.Wnt5a、Dvlをノックダウンした条件でのRac1、RhoA活性の評価
Wnt5a、DvlのノックダウンによりRac1活性は抑制されるのに対して、RhoA活性は上昇している。
図3.FRETによるRac1、RhoA活性局在の評価
極性化したIEC6細胞において、Rac1の活性が基底部に相当する領域に強く、逆にRhoAの活性は頂端部に強い。
図4.極性化した上皮細胞におけるDvl2の局在
IEC6細胞にGFP-Dvl2を発現させると、F-actin cap以外の細胞膜に優位に局在する。
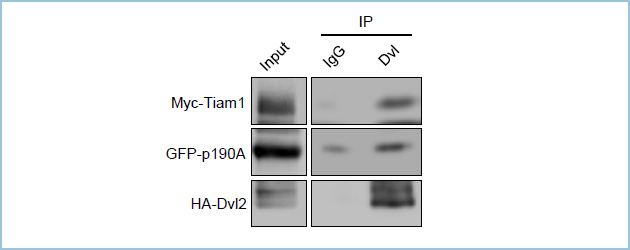
図5.Dvl2とTiam1、p190RhoGAP-Aの複合体形成
HEK293T細胞において、Myc-Tiam1、GFP-p190RhoGAP-A (GFP-p190A)、HA-Dvl2は複合体を形成する。
さらに私共は、IEC6細胞におけるシスト形成の際の頂底極性の確立にWnt5aシグナルが必要となることも確認しており(図6)、多細胞における極性形成においても共通のメカニズムが存在する可能性を考えています。
図6.IEC6細胞のシスト形成におけるWnt5aシグナルの影響
Wnt5a、Dvlのノックダウンにより、シストの頂端部及び基底部の構築が障害される。
上皮細胞におけるWnt5aシグナルによる内腔形成
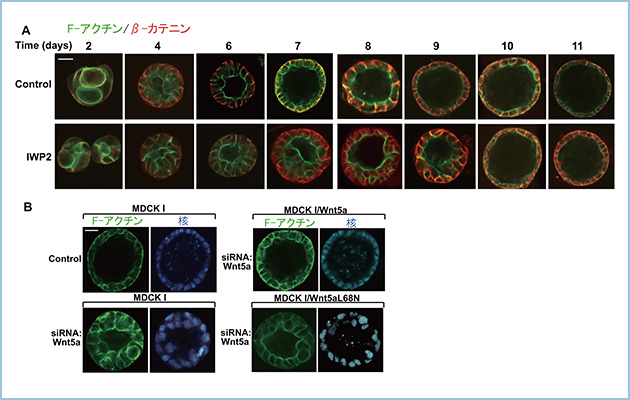
私共はMDCKI細胞のシストにおける内腔形成にWnt5aシグナルが関連することを見出し、Wnt5aが側底側に分泌される生理的役割を解析しました。MDCKI細胞を3Dマトリゲル内で培養すると4日目から内腔形成が始まり、6-7日目にかけて細胞が辺縁で一層に並ぶと共に内腔が大きくなり、8-9日でシストが完成します(図1A)。Porcupine阻害剤のIWP2によってWnt5aの分泌を抑制すると6-7日目において細胞が内腔側に重層し、内腔形成が遅延することから、MDCKI細胞シストの内腔形成にWnt5aシグナルが関与することが示唆されました(図1A)。さらに、Wnt5aの発現抑制による内腔形成の遅延に対して頂端側に分泌されるWnt5aL68Nでは影響しませんが、側底側に分泌される野生型のWnt5aによってレスキューされました。したがって、MDCKI細胞のシストの内腔形成において側底側からWnt5aシグナルを活性化することが重要であると考えられます(図1B)。
図1.MDCKI細胞のシスト形成におけるWnt5aシグナルの影響
A. MDCKI細胞を3Dマトリゲル内で培養すると4日目から内腔形成が始まり、6-7日目にかけて細胞が辺縁で一層に並び内腔が大きくなり、8-9日でシストが完成する。Porcupine阻害剤のIWP2やWnt5aのノックダウンによってWnt5aの分泌を抑制すると6-7日目において細胞が内腔側に重層し、内腔形成が遅延し、10-11日でシストが完成する。
B. Wnt5aの発現抑制による内腔形成の遅延に対して頂端側に分泌されるWnt5aL68Nでは影響しないが、側底側に分泌される野生型のWnt5aによってレスキューされる。
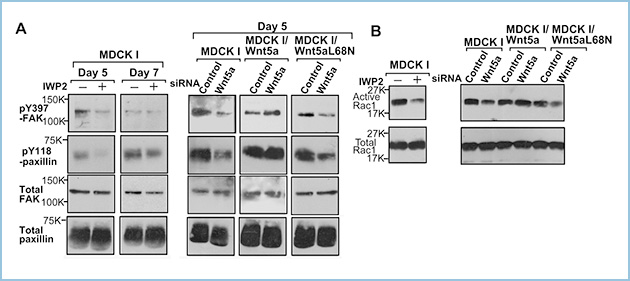
私共はWnt5aシグナルがRac1の活性化を介してIEC6細胞の頂底極性の決定に関与すること、癌細胞株においてWnt5aシグナルが細胞-ECM接着を促進することを報告しています。そこで、極性化されたMDCKI細胞においてWnt5aシグナルがFAKとPaxillinのリン酸化やRac1の活性に対する影響を解析しました。その結果、Wnt5aの発現や分泌を抑制するとFAKやPaxillinのリン酸化、Rac1活性が抑制され、それらの表現型は頂端側に分泌されるWnt5aL68Nでは影響しませんが、側底側に分泌される野生型のWnt5aによってレスキューされました(図2)。したがって、極性化されたMDCKI細胞において側底側からWnt5aシグナルが活性化されることにより、細胞-ECM接着やRac1の活性が促進されることが示唆されました。
図2.MDCKI細胞におけるWnt5aの極性分泌によるFAKとPaxillinのリン酸化やRac1の活性化に対する影響
極性化したMDCKI細胞においてWnt5aの発現や分泌を抑制するとFAKやPaxillinのリン酸化(A)、Rac1活性(B)が抑制される。それらの表現型は側底側に分泌される野生型のWnt5aの発現によってレスキューされるが、頂端側に分泌されるWnt5aL68Nではレスキューされない。
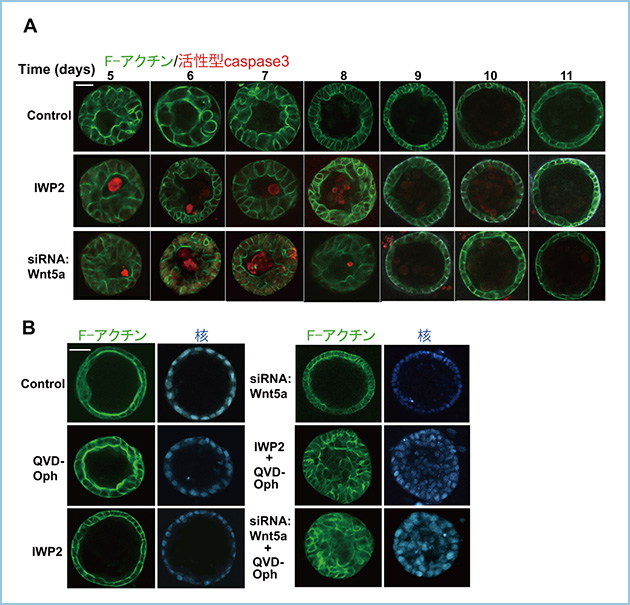
これまでに、細胞接着が抑制されたMDCK細胞のシストの内腔形成(cavitation)においてアポトーシスを利用することが知られています。そこで、Wnt5aシグナルが抑制されたMDCKI細胞におけるシストの内腔形成にアポトーシスが関与するかを解析しました。その結果、コントロール細胞では活性型caspase3によって検出されるアポトーシス細胞は見られませんが、IWP2やノックダウンによってWnt5aの分泌を抑制したMDCKI細胞の内腔ではアポトーシス細胞が検出されました(図3A)。さらに、アポトーシス阻害剤のQVD-Ophを野生型のMDCKI細胞に作用させてもシストの内腔形成に影響しませんが、Wnt5aの発現や分泌を抑制したMDCKI細胞に作用させると内腔形成が抑制されました(図3B)。これらの結果から、Wnt5aシグナルが抑制されると細胞-ECM接着やRac1の活性が抑制され、その結果、アポトーシスによって内腔形成が進行すると考えられます。
極性化されたMDCK細胞においてWnt5a受容体のFz2とRor2受容体が側底側に輸送されることを合わせて考えると、側底側においてWnt5aシグナルが活性化され細胞-ECM接着を促進することにより、内腔形成を促進することが示唆されました。
図3.MDCKI細胞のシスト形成におけるWnt5aシグナル抑制とアポトーシスの関連
A. 野生型のMDCKI細胞のシスト形成において活性型caspase3によって検出されるアポトーシス細胞は検出されないが、Wnt5aの分泌や発現を抑制するとアポトーシス細胞が内腔側に検出され、内腔形成が遅延する。
B. アポトーシス阻害剤のQVD-Ophを野生型のMDCKI細胞に作用させてもシストの内腔形成に影響しないが、Wnt5aの分泌や発現を抑制したMDCKI細胞に作用させると内腔形成が抑制される。
Wnt5aシグナルと上皮形態形成
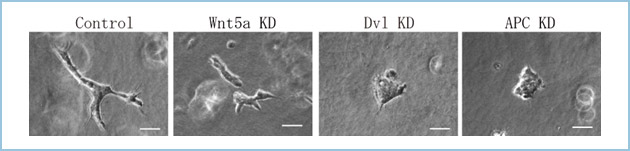
WntはFGFやBMP等と並び、形態形成因子(モルフォゲン)として知られています。アフリカツメガエル、ゼブラフィッシュを用いた発生生物学的解析から、β-カテニン非依存性経路は初期発生段階における原腸形成や神経管形成などの形態形成に関与することが明らかにされています。哺乳動物においてもノックアウトマウスを用いた解析から、β-カテニン非依存性経路が種々の組織、臓器の形態形成を制御することが示唆されています。Wnt5aのノックアウトマウスでは肺や気管の形成不全、小腸の短縮が認められ、心臓の流出路形成に障害をきたします。これらの知見から、β-カテニン非依存性経路は管腔臓器の分枝・形態形成に関与することが示唆されています。これらのマウスのデータに一致して、私達はDvl/APC複合体を介したWnt5aシグナルが三次元培養条件下での上皮細胞の管腔様分枝形態形成にも関与する(図1)ことを明らかにしています。
図1.Wnt5aとDvl/APC複合体の管腔様分枝形態形成への関与
ある種の上皮細胞は細胞外基質内で三次元培養すると、多数の細胞集団からなる管腔様の分枝形態を形成する(Control参照)。私達は、Wnt5a、Dvl、およびAPCをknockdown (KD)した上皮細胞がコラーゲンゲル内三次元培養下での分枝形態形成を抑制することを見出した。スケールバー: 50μm.