A) Molecular mechanisms of signal transduction through cytokine
receptor gp130 in immune response in health and disease: its
implication in autoimmune disease
1) gp130 signals in health and
disease
2) Project
3) Figures for gp130
5) Publication list
6)
OTHER
PROJECTS
1) gp130 signals in health and
disease
Sixteen years have passed since interleukin 6 (IL-6) was cloned in
1986 ( see a review article by
Hirano, Interleukin 6 and its receptor: Ten years later,
International Reviews of Immunology, 16:249-284, 1998). During
the last decade, many findings were made concerning the structure and
function of IL-6 and its receptor, and the role of IL-6 in a variety
of diseases.
Many cytokines possess a similar helical structure. The IL-6
receptor and many other cytokine receptors are also structurally
similar and constitute the cytokine receptor super family. In
addition, cytokine receptor subunits are shared among several
cytokine receptors. This sharing of subunits is one of the mechanisms
by which the functional redundancy of cytokine activities occurs. A
typical example is the gp130 subunit, which is shared among the
receptors for IL-6, leukemia inhibitory factor (LIF), ciliary neurotrophic factor (CNTF), oncostatin M (OSM), IL-11, and
cardiotrophin-1 (CT-1). Studies on the
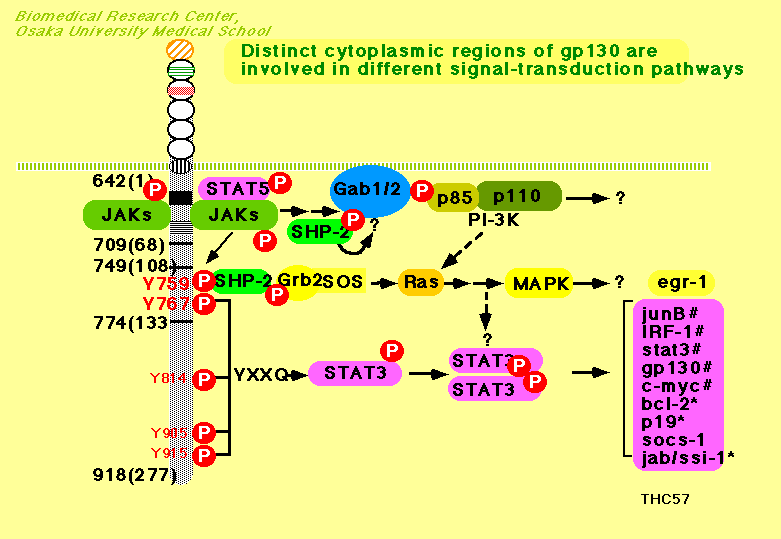
signal transduction of IFNs have shown that novel tyrosine kinases,
JAKs (Janus
kinases), and transcriptional factors, STATs (signal transducer and activator of
transcription), play a major role in signal transduction
through the receptors for a variety of cytokines and hormones.
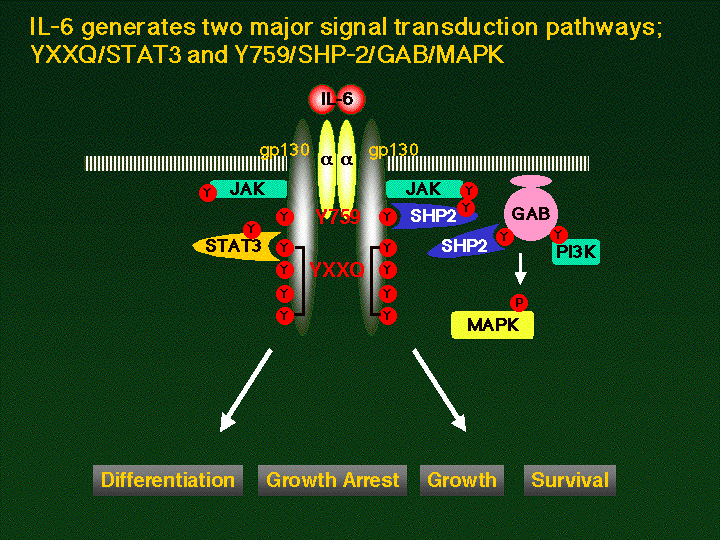
The binding of IL-6 to a chain (CD126)
resulted in the formation of hexametric complex containing two
molecules of each component, IL-6, a chain
and gp130, followed by the activation of Janus kinase (JAK) family
protein tyrosine kinase(s) and tyrosine phosphorylation of various
cellular proteins including gp130 itself. Of the JAK family kinases,
Jak1, Jak2 and Tyk2 constitutively associate with gp130 or gp130-
related LIF receptor and are activated by the IL-6 family of
cytokines. The activated tyrosine kinases, in turn, phosphorylate and
activate the signal transducer and activator of transcription (Stat)
family proteins, especially STAT3 and Stat1 for IL-6 depending on the
phosphorylated YXXQ motif on gp130. Gp130 also has a YXXV motif,
which is recognized by SHP-2 (also referred as PTP1D, SHPTP-2, PTP2C,
and Syp), a member of the phosphotyrosine phosphatase family
containing two SH2 domains, and upon ligand binding SHP-2 has been
shown to be tyrosine phosphorylated in a manner that depends on the
second tyrosine of gp130. SHP-2 has a higher homology with the
Drosophila phosphotyrosine phosphatase, Corkscrew(CSW), which acts
downstream of the Drosophila tyrosine kinase, Torso. Like òCSW, SHP-2
has been shown to act downstream of tyrosine kinases such as the
epidermal growth factor receptor, the fibroblast growth factor
receptor, and the insulin receptor.
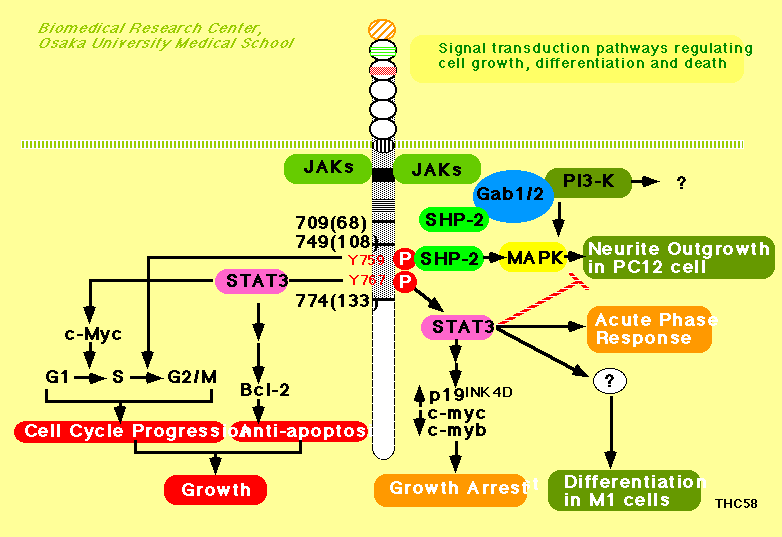
However, the physiological roles of STAT pathway or SHP-2-mediated
pathway through gp130 are largely unknown. To understand the
molecular mechanisms which determine the cell fate through gp130, we
constructed a chimeric receptor consisting of extracelluar domain of
G-CSF receptor or growth hormone receptor and intracellular domain of
gp130. We introduced a series of mutated chimeric receptors into M1
leukemic cells in which IL-6 induces growth arrest and macrophage
differentiation and pro-B cell line, BAF/B03 cells in which IL-6
induces growth. By using the full-length cytoplasmic domain and
mutants with progressive carboxyterminal deletions, we identified the
first 133 amino acid residues of gp130 as the minimal region
necessary and sufficient for both growth arrest and differentiation
signal in M1 cells and growth signal in BAF/B03 cells
(Yamanaka et al, 1996:
Fukada et al, 1996). Immediate early responses,
such as STAT3 activation, junB, egr1 and IRF1 induction, and myb and
myc repression, were observed in M1 clones with chimeric receptors
containing at least the first 133 amino acids of gp130. The mutants
harboring mutations at tyrosine residues that were responsible for
STAT3 activation, did not differentiate into macrophage, or show the
immediate early responses including down regulation of myb and myc.
Our result showed STAT3 activation tightly linked to growth arrest
and differentiation, so STAT3 may play an essential role to generate
growth arrest and differentiation signal in M1
cell(Yamanaka et al, EMBO J., 1996).
Consistent with these observations, we showed that dominant-negative
forms of STAT3 inhibited both IL-6-induced growth arrest and
macrophage differentiation in the M1
transformants(Nakajima et al, EMBO J., 1996).
Blocking of Stat activation resulted in inhibition of IL-6-induced
repression of c-myb and c-myc. Furthermore, IL-6 enhanced the growth
of M1 cells when STAT3 was suppressed. Thus IL-6 generates both
growth-enhancing signals and growth arrest- and
differentiation-inducing signals at the same time in M1 cells and
STAT3 may be critically involved in the cell decision in response to
gp130 signal. We further tried to clarify growth signal in a pro-B
cell line, BAF/B03 cells. We showed that at least two distinct
signals, cell cycle progression and anti-apoptosis, are required for
gp130-induced cell growth(Fukada et al,, Immunity,
1996). The second tyrosine in the YXXV motif (from the membrane)
of gp130, which was required for the tyrosine phosphorylation of
SHP-2, its association with GRB2, and a MAP kinase activation, was
essential for cell cycle progression but not for anti-apoptosis. On
the other hand, the tyrosine in the YXXQ motifs essential for STAT3
activation was required for bcl-2 induction and anti-apoptosis.
Furthermore, dominant negative STAT3 inhibited anti-apoptosis and
bcl-2 induction. These data demonstrate that two distinct signals,
cell cycle progression and anti-apoptosis, are required for
gp130-induced cell growth and STAT3 is involved in anti-apoptosis.
All results suggest that STAT3 play a key role in gp130-mediated
regulation of cell growth, differentiation and suvival. We further
showed that STAT3 plays a key role in the G1 to S phase cell-cycle
transition induced by the cytokine receptor subunit gp130, through
the upregulation of cyclins D2, D3 and A, and cdc25A, and the
concomitant downregulation of p21 and p27. Furthermore, unexpectedly,
we found that gp130 could induce the expression of p21 when STAT3
activation was suppressed. Such contradictory signals regulating
cell-cycle progression could be simultaneously delivered from
distinct cytoplasmic regions of gp130. We propose an 'orchestrating
model' for cytokine and growth factor action in which contradictory
signals are orchestrated to produce a specific effect in a target
cell.(Fukada
et al, EMBO J. ,1998). Thus, the activation of STAT3 by the
cytokine receptor gp130 is required for both the G1 to S cell cycle
transition and antiapoptosis. To clarify this molecular mechanisms,
we tried to identify the target molecules of Stat3. We found that
Pim-1 and Pim-2 are targets for the gp130-mediated STAT3 signal.
Expression of a kinase-defective Pim-1 mutant attenuated
gp130-mediated cell proliferation. Constitutive expression of Pim-1
together with c-myc, another STAT3 target, fully compensated for loss
of the STAT3-mediated cell cycle progression, antiapoptosis, and
bcl-2 expression. We also identified valosine-containing protein
(VCP) as a target gene for the Pim-1-mediated signal. Expression of a
mutant VCP led cells to undergo apoptosis. These results indicate
that Pim-family proteins play crucial roles in gp130-mediated cell
proliferation and explain the synergy between Pim and c-Myc proteins
in cell proliferation and
lymphomagenesis.(Shirogane
et al, Immunity, 1999)
To further elucidate the in vivo roles of gp130-mediated each of
signal transduction pathway, we generated a series of knockin mouse
lines, in which the cytokine receptor gp130-dependent STAT3 and/or
SHP2 signals were disrupted, by replacing the mouse gp130 gene with
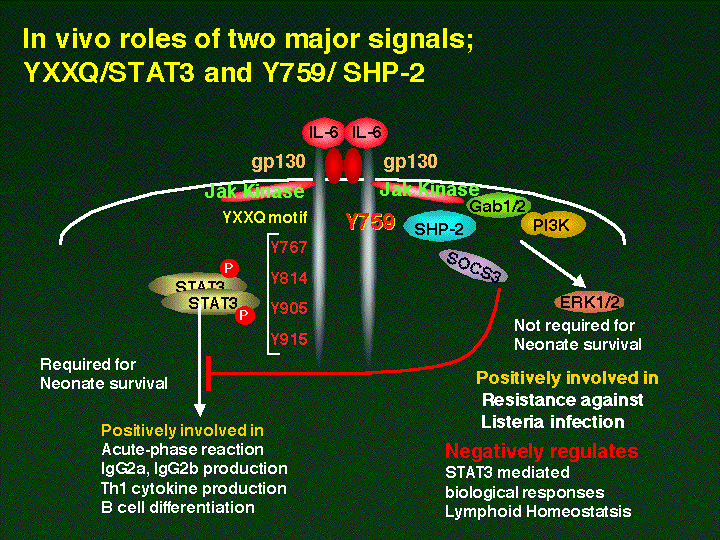
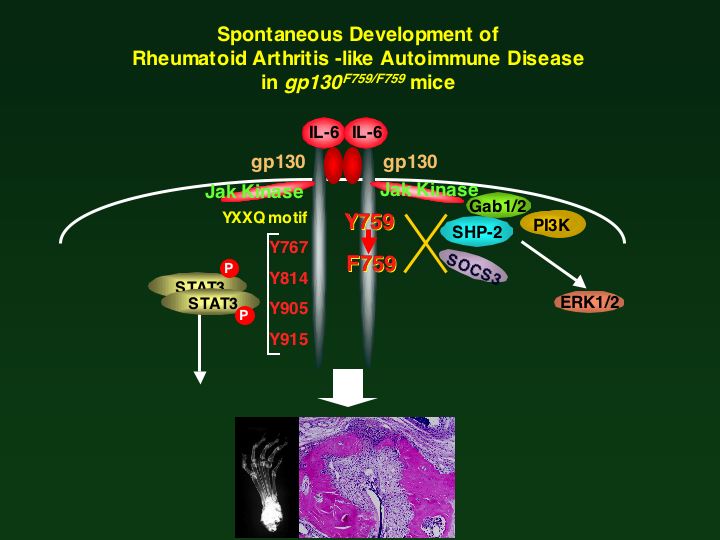
human gp130 mutant cDNAs. The SHP2 signal-deficient mice
(gp130F759/F759 were born normal but displayed splenomegaly and
lymphadenopathy and an enhanced acute phase reaction. In contrast,
the STAT3 signal-deficient mice (gp130FXQ/FXXQ) died perinatally,
like the gp130-deficient mice (gp130D/D). The gp130F759/F759 mice
showed prolonged gp130-induced STAT3 activation, indicating a
negative regulatory role for SHP2. Th1-type cytokine production and
IgG2a and IgG2b production were increased in the gp130F759/F759 mice,
while they were decreased in the gp130FXXQ/FXXQ immune system. Our
results shows that gp130-mediated STAT3 signal plays important roles
in B cell differentiation and Ig production. Furthermore, tyrosine
759 of gp130, SHP-2 binding site, is positively involved in
gp130-mediated MAPK activation, while it negatively regulates
gp130-induced STAT3 activation and Ig production. These results
indicate the balance of the contradictory signals generated through
gp130 determines the final output of the biological activity of IL-6.
(Ohtani
et al , Immunity, 2000). Interesting finding is that the knockin
mice expressing mutant gp130 with a defective in the SHP2 binding
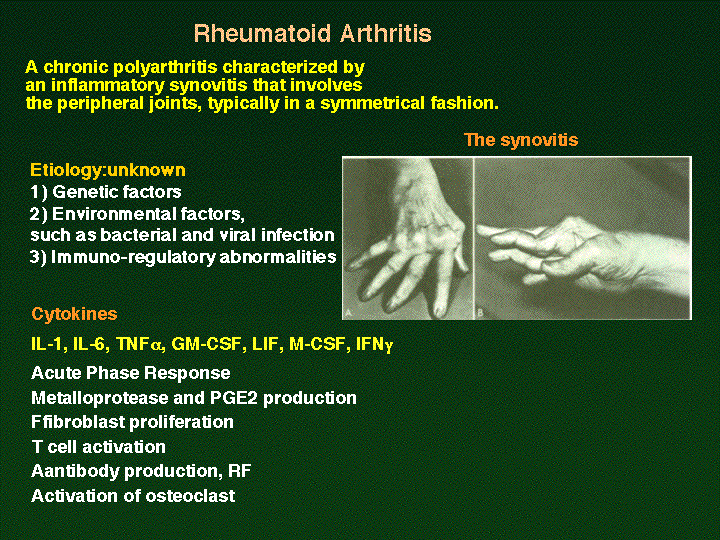
site (F759 mice) spontaneously generate autoimmunity and rheumatoid
arthritis (RA)-like disease as aged (Atsumi, T et al., A point
mutation of Tyr-759 in interleukin 6 family cytokine receptor subunit
gp130 causes autoimmune arthritis. J. Exp Med. 196: 979-990,
2002
(PubMed)).
|

IL-6 induces the differentiation of B cells to antibody
forming cells (Hirano et al,
Nature 324, 73, 1986)
|
IL-6 is a growth factor of myeloma cells
(Kawano et al, Nature 332,
83, 1988). Generation of plasmacytosis and plasmacytoma
in IL-6 transgenic mice
(Suematsu et al, PNAS,86,
7547, 1989; Suematsu et
al,PNAS, 89, 232, 1992)
|
IL-6 induces macrophage differentiation of M1 leukemic
cells (Miyaura et al, FEBS Letters,
234, 17, 1988; Yamanaka
et al, EMBO J, 15, 1557, 1996;
Nakajima et al, EMBO J, 15,
3651, 1996)
|
IL-6 induces neurite outgrowth in PC12 cells
(Satoh et al, Mol. Cell. Biol.,
8, 3546, 1988, Ihara et
al, EMBO J. 17: 5345-5352, 1997.)
|
@
2) Project
1. As described above, the gp130 point mutant knockin mice with a
defective in the gp130-mediated SHP2/Gab/MAPK signal (F759 mice)
spontaneously generate RA-like disease with a frequency of 100 % at
the age of 18 months. We wish to elucidate what cells and what genes
are required for the initiation of the disease and the effecter phase
of RA-like disease in F759 mice. To identify the genetic background
affecting disease development, we will backcross the original F759
mice (129/Bl6 hybrid) to different strains, such as C57BL/6, Balb/c,
DBA/1, 129, C3H/J. To examine whether hematopoietic cells,
non-hematopoietic microenvironment, or both are necessary for the
initiation of RA-like disease in F759 mice, bone marrow transfer
experiment in several combinations will be made. We have already
known that RA-like disease can be transferred to RAG2-/- mice by
spleen cells obtained from the diseased mice. We will examine what
population of spleen cells of the diseased mice can transfer the
disease to RAG2-/- mice. Another important question is what cells and
molecules are required for the initiation of the disease. To answer
this issue we will cross the F759 mice with several kinds of knockout
mice, such as CD4-/-, CD8-/-, mMT-/-, RAG2-/-, IL-6-/-,
stat3-promoter mutant mice so on. We have already made knockin mice
expressing low level of STAT3 due to the point mutation of the stat3
promoter region. We will also try to identify the genes abnormally
expressed in the F759 mice.
2. Elucidation of the effect of environmental factors on
autoimmune diseases in F759 mice. We wish to search the environmental
factors, which affect the onset of RA-like disease in F759 mice. We
will examine the effect of LPS, CFA, bacterial and viral infection
and over expression of IL-6 on the generation of RA in F759 mice. We
have already found that HTLV1-tax expression accelerates the onset of
RA in F759 mice by crossing F759 mice and Tax-transgenic mice in
C57BL/6 genetic background. Under these conditions, RA-like disease
developed as early as 3 months of age. We wish to examine if F759
mutation in hematopoietic lineage cells is enough for the generation
of RA-like disease. If this is the case, we will examine whether the
retrovirus-mediated expression of dominant negative STAT3 in
hematopoietic lineage cells can prevent the disease.@We also take
advantage of Tax-induced acceleration of disease onset to investigate
what populations of hematopoietic cell lineages are required for the
disease initiation and effecter phase for the disease.
3. Elucidation of the immunological state observed in the F759
mice.
To investigate the effect of F759 mutation on thymic negative
selection, we will cross the F759 mice with anti-HY TCR transgenic
mice, and anti-OVA TCR transgenic mice. We also examine if F759
mutaion has any effect on anti-CD3- and superantigen-induced clonal
deletion and activation induced cell death. We have already observed
that activated memory CD4 cells are increased in the diseased mice.
We wish to identify the molecular and immunological mechanisms by
which memory T cells are increased in the F759 mice.
The final goal of our project is the clarification of the
regulatory mechanisms of the immune response from the view point of
signal trasnduction, in particular, cytokine mediated signaling
pathways and establishment of the methods by which we can modurate
and control immune response to prevent or cure autoimmune diseases.
We will continue and extend the experiments planned above.
Furthermore, based on the results obtained from the experiments
performed, we will make several knockout mice to examine if the genes
identified are actually involved in the regulation of the immune
response and autoimmune diseases. We also try to examine the effect
of retrovirus-mediated gene transfer of the dominant negative form of
newly identified genes to bone marrow cells on the generation of the
disease. Moreover, based on the information on the genetic background
affecting the generation of autoimmune disease in F759 mice, we will
try to identify the other genes affecting disease development in F759
mice by simple sequence length polymorphisms (SSLP). Finally, we will
try to search inhibitors for the molecules critically involved in the
immune response, allergic response and autoimmune diseases based on
the results obtained from our research plans.
This project would shed light on the molecular mechanisms of
immune responses, homeostasis of immune system and autoimmune
diseases. Furthermore, F759 mouse is a very useful model mouse to
investigate the molecular mechanisms by which autoimmune disease is
developed and search new drugs to prevent or cure the autoimmune
diseases, such as RA. In summary, this project would greatly
contribute to clarification of the molecular mechanisms of immune
responses and development of new drugs to regulate immune response,
allergic response, and autoimmune diseases.
@
Figures for gp130
@
@
@
Publications:
- Atsumi, T*., K. Ishihara*, D. Kamimura, H. Ikushima, T.
Ohtani, S. Hirota, H. Kobayashi, S-.J. Park, Y. Saeki, Y.
Kitamura, and T. Hirano. (*equal contribution). A point mutation
of Tyr-759 in interleukin 6 family cytokine receptor subunit gp130
causes autoimmune arthritis. J. Exp Med. 196: 979-990, 2002
(PubMed)
- Kamimura, D., D. Fu, Y. Matsuda, T. Atsumi, T. Ohtani, S. J.
Park, K. Ishihara, T. Hirano. Tyrosine 759 of the cytokine
receptor gp130 is involved in Listeria monocytogenes
susceptibility. Genes and Immunity. 3: 136-143, 2002.
(PubMed)
- Narimatsu, M., H. Maeda, S. Itoh, T. Atsumi, T. Ohtani, K.
Nishida, M. Itoh, D. Kamimura, S.-J. Park, K. Mizuno, J. Miyazaki,
M. Hibi, K. Ishihara, K. Nakajima and T. Hirano. Tissue-specific
autoregulation of the stat3 gene and its role in interleukin
6-induced survival signals in T cells. Mol. Cell. Biol.
21:6615-6625, 2001
(PubMed).
- Ohtani, T., K. Ishihara, T. Atsumi, K. Nishida, Y. Kaneko, T.
Miyata , S. Itoh, M. Narimatsu, H. Maeda, T. FukadaÊ, M. Itoh, H.
Okano, M. Hibi and T. Hirano. Dissection of signaling cascades
through gp130 in vivo: Reciprocal roles for STAT3- and
SHP2-mediated signals in immune responses. Immunity, 12,
95-105, 2000.
(PubMed)
(Full
Text in Immunity)
- Shirogane, T., T. Fukada, J.M.M. Muller, D. T. Shima, M. Hibi,
and T. Hirano, Synergistic roles for Pim-1 and c-Myc in
STAT3-mediated cell cycle progression and anti-apoptosis.
Immunity 11, 709-719, 1999.
(PubMed)
(Full
Text in Immunity)
- Kiuchi, N., K. Nakajima, M. Ichiba, T. Fukada, M. Narimatsu,
K. Mizuno, M. Hibi, and T. Hirano. STAT3 is required for the
gp130-mediated full activation of the c-myc gene. J. Exp.
Med. 189: 63-73, 1999
(Abstract)(PubMed)
- Fukada, T., T. Ohtani, Y. Yoshida, T. Shirogane, K. Nishida,
K. Nakajima, M. Hibi, and T. Hirano. STAT3 orchestrates
contradictory signals in cytokine-induced G1 to S cell cycle
transition. EMBO J. 17: 6670-6677,1998
(Abstract)(PubMed)
(Full
Text in EMBO J).
- Ichiba, M., K. Nakajima, Y. Yamanaka, N. Kiuchi and T. Hirano
Autoregulation of the Stat3 gene through cooperation with a CRE
site binding protein. J. Biol. Chem. 273:6132-6138, 1998
(PubMed)
- Ihara, S., K. Nakajima, T. Fukada, M. Hibi, S. Nagata, T.
Hirano, and Y. Fukui. Dual control of neurite outgrowth by STAT3
and MAP kinase in PC12 cells stimulated with interleukin-6.
EMBO J. 16: 5345-5352,
1997(Abstract).
- Two Signals Are Necessary for Cell Proliferation Induced by a
Cytokine Receptor gp130: Involvement of STAT3 in Anti-Apoptosis.
Toshiyuki Fukada, Masahiko Hibi, Yojiro Yamanaka, Mariko
Takahashi-Tezuka, Yoshio Fujitani, Takuya Yamaguchi, Koichi
Nakajima@and Toshio Hirano, Immunity, Vol. 5, 449-460,
1996
(Abstract).(Immunity
on line for Full Text)
- A central role for Stat3 in IL-6-induced regulation of growth
and differentiation in M1 leukemia cells. Nakajima-K; Yamanaka-Y;
Nakae-K; Kojima-H; Ichiba-M; Kiuchi-N; Kitaoka-T; Fukada-T;
Hibi-M; Hirano-T, EMBO-J. 1996 Jul 15; 15(14): 3651-8
(Abstract)
- Differentiation and growth arrest signals are generated
through the cytoplasmic region of gp130 that is essential for
Stat3 activation. Yamanaka-Y; Nakajima-K; Fukada-T; Hibi-M;
Hirano-T, EMBO-J. 1996 Apr 1; 15(7): 1557-65
(Abstract)
- IL-6-inducible complexes on an IL-6 response element of the
junB promoter contain Stat3 and 36 kDa CRE-like site binding
protein(s). Kojima-H; Nakajima-K; Hirano-T, Oncogene. 1996
Feb 1; 12(3): 547-54 (Abstract)
- Transcriptional activation of the IL-6 response element in the
junB promoter is mediated by multiple Stat family proteins.
Fujitani-Y; Nakajima-K; Kojima-H; Nakae-K; Takeda-T; Hirano-T,
Biochem-Biophys-Res-Commun. 1994 Jul 29; 202(2): 1181-7
(Abstract)
- E1A repression of IL-6-induced gene activation by blocking the
assembly of IL-6 response element binding complexes. Takeda-T;
Nakajima-K; Kojima-H; Hirano-T, J-Immunol. 1994 Nov 15;
153(10): 4573-82 (Abstract)
- Identification of a novel interleukin-6 response element
containing an Ets-binding site and a CRE-like site in the junB
promoter. Nakajima-K; Kusafuka-T; Takeda-T; Fujitani-Y; Nakae-K;
Hirano-T, Mol. Cell. Biol. 1993 May; 13(5): 3027-41
(Abstract)
Reviews
- Hirano, T. Cytokines in autoimmune disease and chronic
inflammatory proliferative disease. Editorial for
Cytokine
Growth Factor Rev, 13F297-298,
2002.
(PubMed), RCAI
- Ishihara, K., and Hirano, T. IL-6 in autoimmune disease and
chronic inflammatory proliferative disease.
Cytokine
Growth Factor Rev, 13:357-368, 2002.
(PubMed),
RCAI
- Ishihara, K., and Hirano, T. The molecular basis of the
cell-specificity of cytokine action. BBA, in press,
- Hirano, T. Revival of the autoantibody model in rheumatoid
arthritis. Nature Immunology 3:342-344, 2002,
(PubMed),
- Hibi, M., and Hirano, T. IL-6 receptor. Cytokine
Reference, edited by Oppenheim, J. J., Feldmann, M., Durum, S.
K., Hirano, T., Vilcek, J., Nicola, N.A. Academic Press. Vol. 2,
pp1761-1778. 2001.
(On line)
- Matsuda, T. and Hirano, T. IL-6. Cytokine Reference,
edited by Oppenheim, J. J., Feldmann, M., Durum, S. K., Hirano,
T., Vilcek, J., Nicola, N.A. Academic Press. Vol. 1, pp537-563.
2001.(On
line)
- Hirano, T., and Fukada, T. IL-6 ligand and receptor family.
Cytokine Reference, edited by Oppenheim, J. J., Feldmann,
M., Durum, S. K., Hirano, T., Vilcek, J., Nicola, N.A. Academic
Press. Vol. 1, pp523-535. 2001.
(On line)
- Hirano, T., K. Ishihara, M. Hibi. Roles of STAT3 in mediating
the cell growth, differentiation, and survival signals relayed
through the IL-6 family of cytokine receptors. Oncogene,
19: 2548-2556, 2000.
(PubMed)
- Hirano, T. Molecular basis underlying functional pleiotropy of
cytokines and growth factors. Biochem. Biophys. Res. Comm.,
260, 303-308, 1999
(PubMed).
- Fukada, T., Y. Yoshida, K. Nishida, T. Ohtani, T. Shirogane,
M. Hibi, and T. Hirano. Signaling through gp130: Toward a general
scenario of cytokine action. Growth Factors 17, 81-91,
1999.
(PubMed)
- Hirano, T. Interleukin 6 and its receptor: Ten years later.
Intern. Rev. Immunol., 16:249-284, 1998
(Full
Text)(PubMed)
- Hirano, T., K. Nakajima, and M. Hibi. Signaling mechanisms
through gp130: a model of the cytokine system. Cytokine &
Growth Factor Reviews, 8: 241-252, 1997
(Full Text).
- Hibi, M., Nakajima, K., Hirano, T. IL-6 cytokine family and
signal transduction: a model of the cytokine system. J. Mol.
Med. 74:1-12,
1996.(Abstract)
- Hirano, T., T. Matsuda, and K. Nakajima. Signal transduction
through gp130 that is shared among the receptors for the
interleukin 6 related cytokine subfamily. Stem Cells, 12:
262-277,
1994.(MedLine)