پ@
پi‚Qپjgp130‚©‚ç‚جƒVƒOƒiƒ‹‚جگ¶‘ج“à (in vivo)‚إ‚ج–ًٹ„‚ج‰ً–¾: gp130ƒVƒOƒiƒ‹ˆظڈي‚ة‚و‚ء‚ؤ”ڈا‚·‚éژ©Œب–ئ‰uگ«ٹضگك‰ٹ‚ج”ڈا‹@چ\پ@
پ@
IL-6 induces a variety of funcitons, such as the differentiation of B cells to antibody forming cells, macrophage differentiation of M1 leukemic cells, neurite outgrowth in PC12 cells and growth of myeloma cells.
|
IL-6 induces the differentiation of B cells to antibody forming cells (Hirano et al, Nature 324, 73, 1986) |
IL-6 is a growth factor of myeloma cells (Kawano et al, Nature 332, 83, 1988). Generation of plasmacytosis and plasmacytoma in IL-6 transgenic mice (Suematsu et al, PNAS,86, 7547, 1989; Suematsu et al,PNAS, 89, 232, 1992) |
IL-6 induces macrophage differentiation of M1 leukemic cells (Miyaura et al, FEBS Letters, 234, 17, 1988; Yamanaka et al, EMBO J, 15, 1557, 1996; Nakajima et al, EMBO J, 15, 3651, 1996) |
IL-6 induces neurite outgrowth in PC12 cells (Satoh et al, Mol. Cell. Biol., 8, 3546, 1988) |
پ@
ƒTƒCƒgƒJƒCƒ“‚ح–ئ‰uŒn‚âگ_ŒoŒn‚ ‚é‚¢‚ح”گ¶‚ة‚¨‚¢‚ؤڈd—v‚ب‹@”\‚ً‰ت‚½‚µ‚ؤ‚¢‚éگ¶—ٹˆگ«•ھژq‚إ‚ ‚éپB–{Œ¤‹†‚حƒTƒCƒgƒJƒCƒ“‚ھژَ—e‘ج‚ً‰î‚µ‚ؤ‚ا‚ج‚و‚¤‚ب‹@چ\‚إچ×–E‚ج‘گBپA•ھ‰»پAگ¶‘¶ˆغژ‚ً—U“±‚µ‚¤‚é‚ج‚©‚ً•ھژqƒŒƒxƒ‹‚إ–¾‚ç‚©‚ة‚·‚邱‚ئ‚ً–ع“I‚ئ‚µ‚ؤ‚¢‚éپBgp130‚حپA“–ڈ‰IL-6
(interleukin-6)‚جژَ—e‘ج‚جƒRƒ“ƒ|پ[ƒlƒ“ƒg‚ئ‚µ‚ؤپA“¯’èپEcDNAƒNƒچپ[ƒjƒ“ƒO‚³‚ꂽ‚à‚ج‚إ‚ ‚é‚ھپA‚»‚جŒم‚جŒ¤‹†‚©‚çپAIL-6‚ج‚ف‚ب‚炸LIF
(leukemia Inhibitory Factor)پAOSM (oncostatin M)پACNTF (ciliary
neurotrophic factor)پAIL-11پACT-1
(cardiotrophin-1)‚ئ‚¢‚ء‚½IL-6ƒtƒ@ƒ~ƒٹپ[ƒTƒCƒgƒJƒCƒ“‚جژَ—e‘ج‚ج‹¤’تƒRƒ“ƒ|پ[ƒlƒ“ƒg‚إ‚ ‚邱‚ئ‚ھ–¾‚ç‚©‚ئ‚ب‚ء‚ؤ‚¢‚éپB‚»‚جˆêژںچ\‘¢‚حپA‘¼‚جƒTƒCƒgƒJƒCƒ“ژَ—e‘جپiG-CSFژَ—e‘جپAIL-3/IL-5/GM-CSFژَ—e‘جپAƒGƒٹƒXƒچƒ|ƒGƒ`ƒ“ژَ—e‘جپAژيپX‚جinterleukinژَ—e‘ج“™پjپEƒzƒ‹ƒ‚ƒ“ژَ—e‘جپigrowth
hormoneژَ—e‘جپAprolactinژَ—e‘جپAleptinژَ—e‘ج“™پj‚ئچ×–EٹO—جˆو‚ة•غ‘¶‚³‚ꂽ—جˆوپiWSXWSƒ‚ƒ`پ[ƒt‚¨‚و‚رcysteinژcٹî‚جˆت’uپj‚ًژ‚؟پAIŒ^ƒTƒCƒgƒJƒCƒ“ژَ—e‘جƒXپ[ƒpپ[ƒtƒ@ƒ~ƒٹپ[‚ئŒؤ‚خ‚ê‚ؤ‚¢‚éپBIL-6ƒtƒ@ƒ~ƒٹپ[ƒTƒCƒgƒJƒCƒ“‚حپAgp130‚ً‰î‚µ‚ؤپATچ×–EپEBچ×–E“™‚ج–ئ‰uچ×–EپA‘¢ŒŒچ×–EپAٹجچ×–EپAگ_Œoچ×–E‚ة‘خ‚µ‚ؤپA‘گBپE•ھ‰»پEچ×–Eژ€‚ج—}گ§‚ب‚ا‘½ژي‘½—l‚بƒVƒOƒiƒ‹‚ًچ×–E“à‚ض“`‚¦‚邱‚ئ‚ھ’m‚ç‚ê‚ؤ‚¢‚é‚ھپA‚»‚جƒپƒJƒjƒYƒ€‚ح–¾‚ç‚©‚إ‚ب‚¢پB
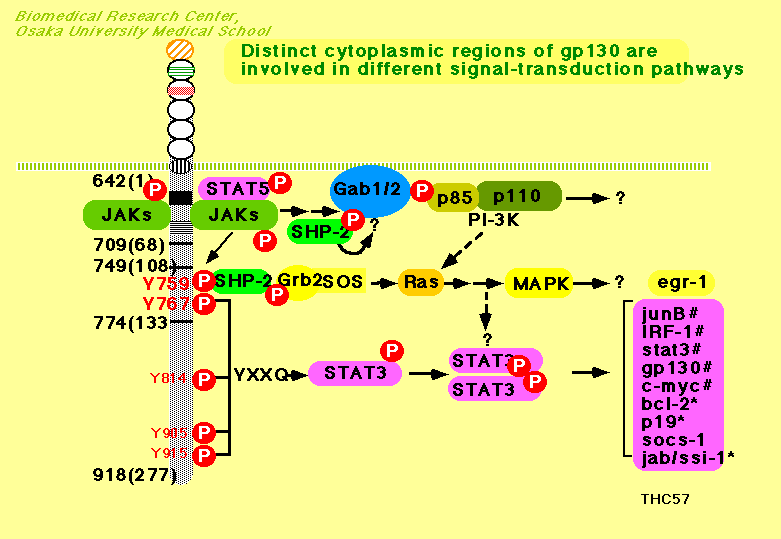
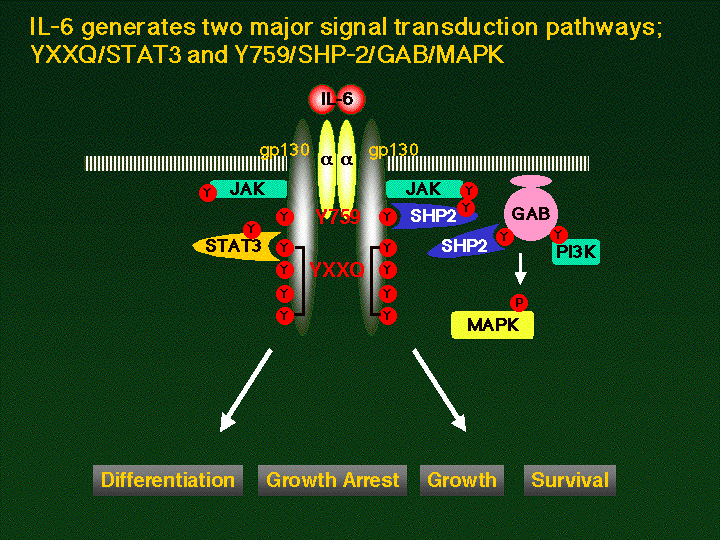
پ@gp130‚ً‰î‚·‚éƒVƒOƒiƒ‹“`’B‚حپAIL-6‚جƒVƒOƒiƒ‹“`’B‚ً’†گS‚ة‰ًگح‚³‚ê‚ؤ‚«‚½پBIL-6‚حپAچ×–E–Œڈم‚ة‘¶چف‚·‚éIL-6ژَ—e‘جa‚ئŒ‹چ‡‚·‚éپB‚³‚ç‚ةپAIL-6ژَ—e‘جa‚حپAgp130‚ئ‰ïچ‡‚·‚é‚ئ‚ئ‚à‚ةgp130‚ا‚¤‚µ‚ج“ٌ—ت‘ج(homodimer)Œ`گ¬‚ً—U“±‚·‚éپBgp130‚جچ×–E“à—جˆو‚ة‚حپA
IŒ^ƒTƒCƒgƒJƒCƒ“ژَ—e‘جƒXپ[ƒpپ[ƒtƒ@ƒ~ƒٹپ[ٹش‚إ•غ‘¶‚³‚ꂽپA—جˆوBox1,
Box2چ\‘¢‚ھ‘¶چف‚µپA‚±‚ج•”•ھ‚ةƒ`ƒچƒVƒ“ƒLƒiپ[ƒ[JAK
(Janus kinase)ƒtƒ@ƒ~ƒٹپ[‚ة‘®‚·‚éJAK1, JAK2,
TYK2‚ھچ\گ¬“I‚ة‰ïچ‡‚·‚邱‚ئ‚ھ–¾‚ç‚©‚ئ‚ب‚ء‚ؤ‚¢‚éپBژhŒƒ‚ة‚و‚èپAgp130‚ھ“ٌ—ت‘ج‚ًŒ`گ¬‚·‚é‚ئ‚ئ‚à‚ةپAgp130‚ة‰ïچ‡‚·‚éJAK‚à‘ٹŒفگع‹ك‚µپAJAK‚ا‚¤‚µ‚ًƒ`ƒچƒVƒ“ƒٹƒ“ژ_‰»‚·‚邱‚ئ‚ة‚و‚èٹˆگ«‰»‚·‚é‚à‚ج‚ئچl‚¦‚ç‚ê‚ؤ‚¢‚éپB‚³‚ç‚ةپAٹˆگ«‰»‚³‚ꂽJAK‚حپAJAK‚ًƒٹƒ“ژ_‰»‚·‚邾‚¯‚إ‚ب‚پAgp130چ×–E“à—جˆو‘¶چف‚·‚éƒ`ƒچƒVƒ“ژcٹ‚و‚رپAژيپX‚جƒVƒOƒiƒ‹“`’B•ھژq‚ًƒٹƒ“ژ_‰»‚µپAٹˆگ«‰»‚·‚邱‚ئ‚ھ’m‚ç‚ê‚ؤ‚¢‚éپB‚±‚ê‚çƒVƒOƒiƒ‹“`’B•ھژq‚ج“àپA“]ژتˆِژqSTAT3
(signal tranducer and activator of transcription
3)‚حپA•ھژq“à‚ةپA“ءˆظ“Iƒٹƒ“ژ_‰»ƒ`ƒچƒVƒ“چ\‘¢‚ً”Fژ¯‚·‚éSH2 (src
homology
2)ƒhƒپƒCƒ“‚ً—L‚µپAgp130چ×–E“à—جˆوƒٹƒ“ژ_‰»ƒ`ƒچƒVƒ“‚ً“ءˆظ“I‚ة”Fژ¯‚µپAgp130ڈم‚ة‰^‚خ‚ê‚ؤ‚‚é‚ئچl‚¦‚ç‚êپAJAK‚ة‚و‚èƒ`ƒچƒVƒ“ƒٹƒ“ژ_‰»‚³‚ê‚éپBƒ`ƒچƒVƒ“ƒٹƒ“ژ_‰»‚³‚ꂽSTAT3‚حپAژ©گg‚جSH2ƒhƒپƒCƒ“‚ً‰î‚µ‚ؤSTAT3“ٌ—ت‘ج(homodimer)‚ ‚é‚¢‚حSTAT1‚ئ‚ج“ٌ—ت‘ج(heterodimer)‚ًŒ`گ¬‚µپAٹj“à‚ضˆعچs‚µپA“ءˆظ“IDNA”z—ٌ‚ً”Fژ¯‚µ‚ؤŒ‹چ‡‚µپA‘½‚‚جˆâ“`ژq‚ج“]ژت‚ًگ§Œن‚µ‚ؤ‚¢‚邱‚ئ‚ھ’m‚ç‚ê‚ؤ‚¢‚éپB‚ـ‚½پAƒٹƒ“ژ_‰»ƒ`ƒچƒVƒ““ءˆظ“I’Eƒٹƒ“ژ_‰»چy‘f(phosphotyrosine
phosphatase)‚جˆê‚آ‚إ‚ ‚éSHP-2‚àپASTAT3“¯—lپA•ھژq“à‚ةSH2ƒhƒپƒCƒ“‚ً—L‚µپAgp130‚جƒ`ƒچƒVƒ“ƒٹƒ“ژ_‰»‚ئ‚ئ‚à‚ةgp130ڈم‚ض‰^‚خ‚êپAJAK‚ة‚و‚èƒ`ƒچƒVƒ“ƒٹƒ“ژ_‰»‚³‚ê‚é‚ئچl‚¦‚ç‚ê‚ؤ‚¢‚éپBgp130‚حپAچ×–E“à—جˆو‚ة‚UŒآ‚جƒ`ƒچƒVƒ“ژcٹî‚ًژ‚ء‚ؤ‚¢‚é‚ھپA‚»‚ج“àپAچ×–E–Œ—جˆو‚©‚çگ”‚¦‚ؤ‚Q”ش–ع‚جƒ`ƒچƒVƒ“(Y2,
Y759)‚حپA‚»‚جژü•سƒAƒ~ƒmژ_”z—ٌپiYSTVپj‚ھپASHP-2‚جSH2ƒhƒپƒCƒ“‚ة‚و‚è”Fژ¯‚³‚ê‚é”z—ٌ‚إ‚ ‚邱‚ئپA‚±‚جƒ`ƒچƒVƒ“‚جƒٹƒ“ژ_‰»‚ھSHP-2‚جƒٹƒ“ژ_‰»‚ة•Kگ{‚إ‚ ‚邱‚ئ‚ھ–¾‚ç‚©‚ئ‚ب‚ء‚ؤ‚¢‚éپB‚ـ‚½پA‚R”ش–ع‚©‚ç‚U”ش–ع‚جƒ`ƒچƒVƒ“ژcٹî‚حپA‚·‚ׂؤƒ`ƒچƒVƒ“‚جC––’[‘¤‚R”ش–ع‚جˆت’u‚ةƒOƒ‹ƒ^ƒ~ƒ“‚ً—L‚µ‚ؤ‚¨‚èپiYXXQƒ‚ƒ`پ[ƒtپjپA‚±‚جƒ‚ƒ`پ[ƒt‚ھˆê‚آ‚إ‚à‘¶چف‚·‚ê‚خپASTAT3‚جٹˆگ«‰»‚ھ—U“±‚³‚ê‚邱‚ئ‚ھ–¾‚ç‚©‚ئ‚ب‚ء‚ؤ‚¢‚éپB
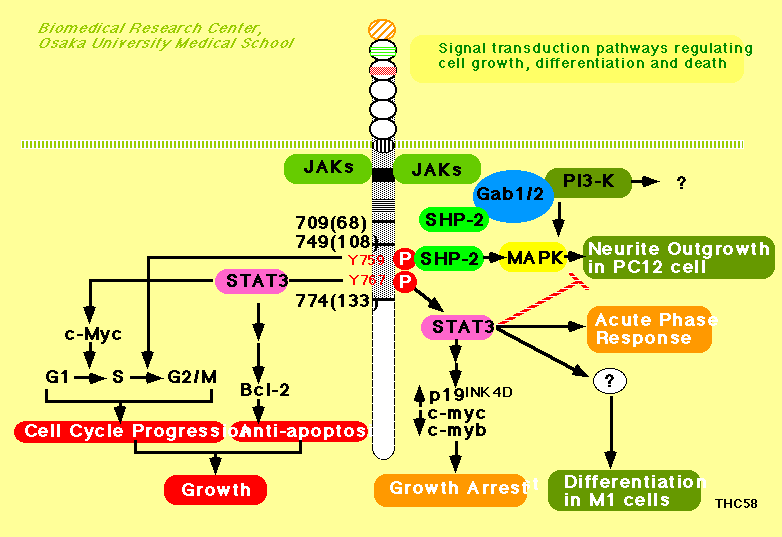
‰نپX‚حپAgp130‚©‚ç‚جƒVƒOƒiƒ‹“`’B‚ً‰ًگح‚·‚邽‚كپAgp130‚جچ×–EٹO—جˆو‚ًGrowth
hormoneپiGHپj‚ ‚é‚¢‚حG-CSFژَ—e‘ج‚ة’u‚«ٹ·‚¦‚½ƒLƒپƒ‰ژَ—e‘ج‚¨‚و‚ر‚»‚جچ×–E“à—جˆو‚ةژيپX‚ج•دˆظ‚ً“±“ü‚µ‚½ژَ—e‘ج‚ًچىگ¬‚µپAژيپX‚جچ×–E‚ةŒ`ژ؟“±“ü‚µپAژيپX‚جƒVƒOƒiƒ‹“`’B‚ة•K—v‚بgp130‚ج—جˆو‚ًŒں“¢‚µ‚ؤ‚«‚½پB‚»‚جŒ‹‰تپAƒ}ƒEƒX”’ŒŒ•aچ×–Eٹ”M1چ×–E‚جچ×–E‘گB’âژ~پEƒ}ƒNƒچƒtƒ@پ[ƒW‚ض‚ج•ھ‰»‚ة‚حپAYXXQƒ‚ƒ`پ[ƒt‚©‚ç‚جSTAT3‚جٹˆگ«‰»‚ھ•Kگ{‚إ‚ ‚邱‚ئپi
EMBO J. 15:3651-3658, 1996.
(Abstract)(PubMed)پA
EMBO J. 15:1557-1565, 1996.
(Abstract)
(PubMed)پAƒ}ƒEƒXB‘O‹ىچ×–Eٹ”BAF-B03‚جgp130ˆث‘¶گ«‚جچ×–E‘گB‚ة‚حپA‚Q”ش–ع‚جƒ`ƒچƒVƒ“ژcٹî‚©‚çپiSHP-2‚ً‰î‚µ‚ؤ‚¢‚é‰آ”\گ«‚ھ‹‚¢پj‚جچ×–E•ھ—ô‘£گi‚جƒVƒOƒiƒ‹‚ئپAYXXQƒ‚ƒ`پ[ƒt‚©‚çSTAT3‰î‚µ‚½چ×–Eژ€—}گ§‚جƒVƒOƒiƒ‹‚ج“ٌ‚آ‚جƒVƒOƒiƒ‹‚ھ•K—v‚إ‚ ‚邱‚ئ‚ًŒ©‚¢ڈo‚µ‚½پiImmunity
5: 449-460, 1996. (Abstract)
(PubMed)(Immunity
on line for Full
TEXT)پB‚ـ‚½STAT3‚ج•W“Iˆâ“`ژq‚ئ‚µ‚ؤc-myc‚ئpim1/2‚ً–¾‚ç‚©‚ة‚µ‚½پiJ.
Exp. Med. 189: 63-73, 1999
(Abstract)(PubMed)پAImmunity
11, 709-719, 1999.
(PubMed)
(Full
Text in
Immunity)پjپBˆê•ûپAgp130ژhŒƒ‚ة‚و‚éPC12چ×–E‚جگ_Œoچ×–E‚ض‚ج•ھ‰»‚ة‚ح‚Q”ش–ع‚جƒ`ƒچƒVƒ“(Y2,
Y759)‚ً‰î‚·‚éMAPK‚جٹˆگ«‰»‚ھڈd—v‚إ‚ ‚èپA‚R”ش–ع‚¢‚©‚جƒ`ƒچƒVƒ“پiY3-Y6پj‚ً‰î‚·‚éSTAT3‚جٹˆگ«‰»‚ح—}گ§“I‚ةچى—p‚·‚邱‚ئ‚ً–¾‚ç‚©‚ة‚µ‚½پiEMBO
J. 17: 5345-5352,
1997(Abstract).(PubMed)پB‚³‚ç‚ةپASHP-2‚¨‚و‚رSTAT3‚جƒVƒOƒiƒ‹ˆبٹO‚ةپAJAK‚©‚ç’¼گعSTAT5پiSTAT3“¯—lSTATƒtƒ@ƒ~ƒٹپ[‚à‘®‚·‚éپj‚ھٹˆگ«‰»‚³‚ê‚邱‚ئپiOncogene,
14: 751-761, 1997.
(Abstract)(PubMed)پAJAKˆبٹO‚ة‚àTecŒ^‚جƒ`ƒچƒVƒ“ƒLƒiپ[ƒ[پiTec/Btkپj‚ھٹˆگ«‰»‚³‚êپA‚»‚ج‰؛—¬‚ةPI-3
kinase‚âƒAƒ_ƒvƒ^پ[•ھژqVav‚ھ‘¶چف‚·‚邱‚ئ‚ًŒ©‚¢ڈo‚µ‚½پiOncogene,
14: 2273-2282, 1997
(Abstract)(PubMed)پB‚³‚ç‚ةپAgp130‚©‚ç‚جگV‹K‚جƒVƒOƒiƒ‹ŒoکH‚ئ‚µ‚ؤپAƒAƒ_ƒvƒ^پ[•ھژqGab1‚ھgp130‚جƒ`ƒچƒVƒ“ƒٹƒ“ژ_‰»”ٌˆث‘¶گ«‚ةJAK‚ة‚و‚èƒٹƒ“ژ_‰»‚³‚ê‚邱‚ئپBGab1‚ھپAgp130ژhŒƒˆث‘¶گ«‚ةSHP-2‚¨‚و‚رPI-3ƒLƒiپ[ƒ[‚ئ‰ïچ‡‚·‚邱‚ئپAGab1‚جƒVƒOƒiƒ‹“`’B‰؛—¬‚ةMAPƒLƒiپ[ƒ[ERK‚ھ‘¶چف‚µ‚ؤ‚¢‚邱‚ئ‚©‚çپAGab1‚ھSHP-2‚©‚çERK‚ض‚جƒVƒOƒiƒ‹“`’B‚ةڈd—v‚ب–ًٹ„‚ً‰ت‚½‚µ‚ؤ‚¢‚é‰آ”\گ«‚ً–¾‚ç‚©‚ة‚µ‚ؤ‚«‚½(Mol.
Cell. Biol. 18:4109-4117, 1998.
(PubMed)(Full
Text in
MCB)پjپB‚³‚ç‚ةGab2‚ًƒNƒچپ[ƒjƒ“ƒO‚µپAGabƒtƒ@ƒ~ƒٹپ[‚ھژيپX‚جƒTƒCƒgƒJƒCƒ“‚â‘گBˆِژqپAچRŒ´ژَ—e‘ج‚ً‰î‚·‚éƒVƒOƒiƒ‹“`’B‚ةٹض—^‚µ‚ؤ‚¢‚邱‚ئ‚ً–¾‚ç‚©‚ة‚µ‚½پiBlood,
93:1809-1816,
1999.(PubMed)).پ@‚ـ‚½Gab1ƒmƒbƒNƒAƒEƒgƒ}ƒEƒX‚ًچىگ¬‚µپAGab1‚ھƒTƒCƒgƒJƒCƒ“‚©‚çMAPK‚ض‚جƒVƒOƒiƒ‹“`’B‚ة•Kگ{‚إ‚ ‚邱‚ئ‚ً–¾‚ç‚©‚ة‚·‚é‚ئ‚ئ‚à‚ةپAگS‘ں‚â‘ظ”ص‚جŒ`گ¬‚ة•Kگ{‚ج–ًٹ„‚ً’S‚ء‚ؤ‚¢‚邱‚ئ‚ً–¾‚ç‚©‚ة‚µ‚½پiMol.
Cell. Biol. 20, 3695-3704,
2000.(PubMed)(Abstract)(Full
Text in
MCBپjپB‚³‚ç‚ة‰نپX‚حگ¶‘ج“à‚ة‚¨‚¯‚égp130ƒVƒOƒiƒ‹‚ج–ًٹ„‚ً–¾‚ç‚©‚ة‚·‚邽‚ك‚ةپAgp130‚ً‰î‚·‚éSTAT3‚ ‚é‚¢‚حSHP2ƒVƒOƒiƒ‹‚ج‚ف‚ً“ءˆظ“I‚ةŒ‡ژ¸‚µ‚½gp130پA‚·‚ب‚ي‚؟ƒVƒOƒiƒ‹“ءˆظ“I•دˆظgp130‚ً”Œ»‚µ‚ؤ‚¢‚éƒmƒbƒNƒCƒ“ƒ}ƒEƒX‚ًژ÷—§‚µ‚½پiImmunity,
12, 95-105, 2000.
(PubMed)
(Full
Text in
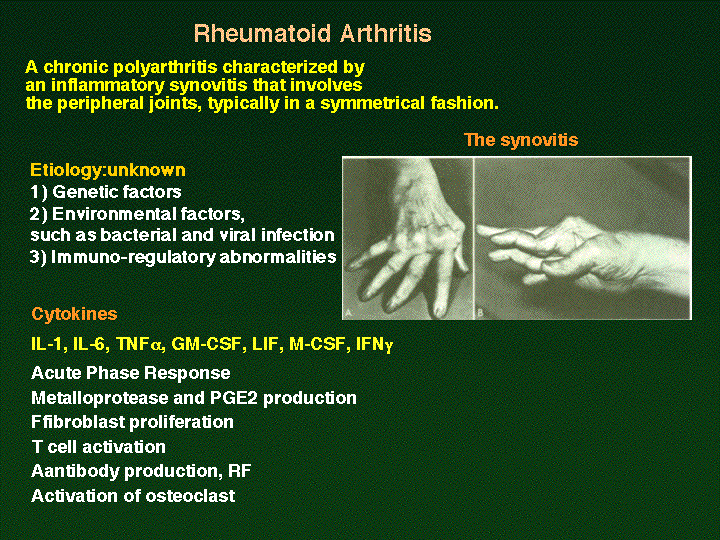
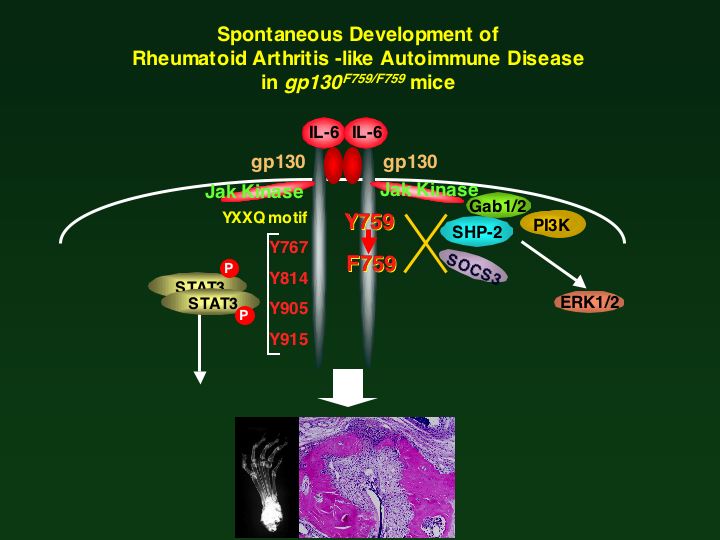
Immunity)پjپB‚³‚ç‚ةڈd—v‚ب“_‚حSHP2ƒTƒCƒg‚ة“_•دˆظ‚ً“±“ü‚µ‚½gp130‚ً”Œ»‚µ‚ؤ‚¢‚éƒmƒbƒNƒCƒ“ƒ}ƒEƒX‚حگ¶Œم1پ|1”N”¼Œم‚ة‚P‚O‚Oپ“ٹضگكƒٹƒEƒ}ƒ`—lژ©Œب–ئ‰uژ¾ٹ³‚ً”ڈا‚·‚邱‚ئ‚ھ–¾‚ç‚©‚ة‚ب‚ء‚½(Atsumi,
T et al., A point mutation of Tyr-759 in interleukin 6 family
cytokine receptor subunit gp130 causes autoimmune arthritis. J.
Exp Med. 196: 979-990, 2002
(PubMed))پB‚±‚جژ–ژہ‚و‚èپAƒCƒ“ƒ^پ[ƒچƒCƒLƒ“‚Uƒtƒ@ƒ~ƒٹپ[ƒTƒCƒgƒJƒCƒ“‚جƒVƒOƒiƒ‹ˆظڈي‚ة‚و‚ء‚ؤٹضگكƒٹƒEƒ}ƒ`—lژ©Œب–ئ‰uژ¾ٹ³‚ھ”ڈا‚µ‚¤‚邱‚ئ‚ھڈ‰‚ك‚ؤڈط–¾‚³‚ꂽپBچ،Œمژ©Œب–ئ‰uژ¾ٹ³‚ج”ڈا‹@چ\‚ً‰ً–¾‚·‚邽‚ك‚ة”ٌڈي‚ة—L—p‚بƒ‚ƒfƒ‹ƒ}ƒEƒX‚ة‚ب‚éپB
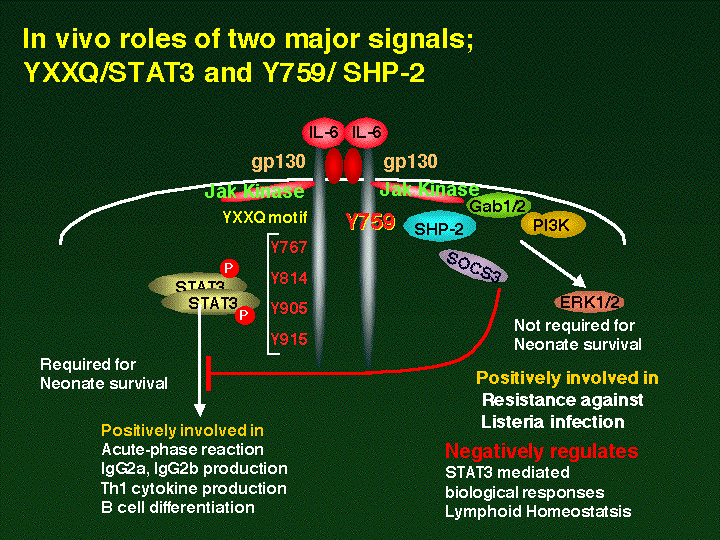
ˆبڈم‚ج—l‚ةپAgp130‚ً‰î‚·‚éƒVƒOƒiƒ‹‚ح‘ه‚«‚پA(i) JAK‚©‚ç‚جgp130‚جƒٹƒ“ژ_‰»‚ً‰î‚µ‚ب‚¢’¼گع‚جƒVƒOƒiƒ‹پA(ii) gp130چ×–E“àƒ`ƒچƒVƒ“759‚جƒٹƒ“ژ_‰»ˆث‘¶گ«‚ةƒٹƒ“ژ_‰»‚³‚ê‚éSHP-2‚ً‰î‚·‚éƒVƒOƒiƒ‹پA(iii) gp130C––’[YXXQƒ‚ƒ`پ[ƒt‚جƒ`ƒچƒVƒ“ƒٹƒ“ژ_‰»ˆث‘¶گ«‚ةƒٹƒ“ژ_‰»ٹˆگ«‰»‚³‚ê‚éSTAT3‚ً‰î‚·‚éƒVƒOƒiƒ‹پA‚ج‚R‚آ‚ة•ھ‚¯‚邱‚ئ‚ھ‚إ‚«‚éپB‚±‚ê‚ç‰نپX‚ھ‚P‚O”N‚©‚¯‚ؤ–¾‚ç‚©‚ة‚µ‚ؤ‚«‚½Œ¤‹†گ¬‰ت‚ً‚à‚ئ‚ةپAGabƒtƒ@ƒ~ƒٹپ[‚ً‰î‚·‚é–ئ‰uپAٹàپAƒTƒCƒgƒJƒCƒ“ƒVƒOƒiƒ‹‚جŒ¤‹†پAgp130ƒVƒOƒiƒ‹ˆظڈي‚ة‚و‚ء‚ؤ”ڈا‚·‚éژ©Œب–ئ‰uژ¾ٹ³‚ج”ڈا‹@چ\‚ج–ئ‰uچ×–EƒŒƒxƒ‹پA•ھژqچ×–EƒŒƒxƒ‹‚إ‚جŒ¤‹†‚ًڈW’†‚µ‚ؤچs‚¤پB
پ@
پ@
پ@‰نپX‚حگ¶‘ج“à‚ة‚¨‚¯‚égp130ƒVƒOƒiƒ‹‚ج–ًٹ„‚ً–¾‚ç‚©‚ة‚·‚邽‚ك‚ةپAƒVƒOƒiƒ‹“ءˆظ“I•دˆظgp130‚ً”Œ»‚µ‚ؤ‚¢‚éƒmƒbƒNƒCƒ“ƒ}ƒEƒX‚ًژ÷—§‚µ‚½پiImmunity, 12, 95-105, 2000. (PubMed) (Full Text in Immunity)پjپBƒqƒggp130‚جƒ`ƒچƒVƒ“759‚¨‚و‚رYXXQƒ‚ƒ`پ[ƒt‚ًژ‚ء‚½ƒ`ƒچƒVƒ“767, 824, 905, 915‚ًƒtƒFƒjپ[ƒ‹ƒAƒ‰ƒjƒ“‚ة•دٹ·‚µ‚½•دˆظ‘ج(F2, F3-6)‚حپA‚»‚ꂼ‚êSHP-2پESTAT3‚ً‰î‚·‚éƒVƒOƒiƒ‹‚ً“`’B‚·‚邱‚ئ‚ھ‚إ‚«‚ب‚¢پiƒVƒOƒiƒ‹•دˆظ‘جپjپB‚±‚ê‚ç•دˆظ‘ج‹y‚ر–ىگ«Œ^cDNA‚جچ×–E–ŒپEچ×–E“à—جˆو‚ًٹـ‚ق’f•ذ‚ًƒ}ƒEƒXESچ×–E‚جgp130گُگF‘جDNA‚ج“¯‚¶•”•ھ‚ة‘ٹ“¯‘g‚فٹ·‚¦–@‚ة‚و‚è‘}“ü‚·‚éپiƒmƒbƒNƒCƒ“ƒ}ƒEƒXپjپB‚±‚جESچ×–E‚ة‚¨‚¢‚ؤ‚حپA‘g‚فچ‚ـ‚ꂽˆâ“`ژqڈم‚إ‚حچ×–EٹO‚ًƒ}ƒEƒXgp130پAچ×–E–ŒپEچ×–E“à—جˆو‚ًƒqƒggp130‹y‚ر‚»‚ج•دˆظ‘ج‚ة’uٹ·‚³‚ꂽƒ}ƒEƒXپ[ƒqƒgƒLƒپƒ‰gp130‚ً”Œ»‚·‚éپB‚±‚جESچ×–E‚ًمَ–E—‘‚ة’چ“ü‚µƒ}ƒEƒXژq‹{‘ج‚ة–ك‚µپAƒLƒپƒ‰ƒ}ƒEƒX‚ًچىگ¬‚·‚éپBŒً”z‚ة‚و‚èچإڈI“I‚ة‘ٹ“¯گُگF‘ج—¼•û‚جgp130ˆâ“`ژq‚ة•دˆظ‚ً—L‚·‚éƒ}ƒEƒX‚ًچىگ¬‚µ‚½پB‚»‚جŒ‹‰تپAgp130‚ً‰î‚·‚éSTAT3ƒVƒOƒiƒ‹‚حچR‘جژYگ¶پABچ×–E•ھ‰»“™‚ة•K—v‚إ‚ ‚邱‚ئپASHP2ƒVƒOƒiƒ‹‚حMAPK‚جٹˆگ«‰»‚ة‚ح•Kگ{‚¾‚ھپASTAT3ƒVƒOƒiƒ‹‚ة‚ح•‰‚جچى—p‚ً‚µ‚ؤ‚¢‚邱‚ئ‚ً–¾‚ç‚©‚ة‚µ‚½پBچإ‹كSHP2ƒVƒOƒiƒ‹‚ًŒ‡‘¹‚µ‚½gp130”Œ»ƒ}ƒEƒX‚ة‚¨‚¢‚ؤگ¶Œم‚P”Nپ[‚P”N”¼Œo‰ك‚·‚é‚ئ‚ظ‚ع‚P‚O‚Oپ“‚جŒآ‘ج‚ة‚¨‚¢‚ؤژ©Œب–ئ‰uگ«‚جٹضگك‰ٹ‚ھ”ڈا‚·‚邱‚ئ‚ھ–¾‚ç‚©‚ة‚ب‚ء‚½پB‚±‚جٹضگك‰ٹ‚حƒqƒg‚ج–گ«ٹضگكƒٹƒEƒ}ƒ`پiRAپj‚ةچ“ژ—‚µ‚ؤ‚¨‚èپARA‚ج”ڈا‹@چ\‚ًŒ¤‹†‚·‚éڈم‚إ‘ه•د‹Mڈd‚ب“®•¨ƒ‚ƒfƒ‹‚إ‚ ‚邱‚ئ‚ھ–¾‚ç‚©‚ة‚ب‚ء‚½پBŒ»چف‚³‚ç‚ة–ئ‰uٹw“I‚ةڈعچׂب‰ًگح‚ًچs‚ء‚ؤ‚¢‚éپB‚ـ‚½‚ب‚؛gp130‚ً‰î‚·‚éƒVƒOƒiƒ‹ˆظڈي‚ھژ©Œب–ئ‰uژ¾ٹ³‚ً”ڈا‚·‚é‚ج‚©‚ئ‚¢‚¤“_‚ةڈإ“_‚ًˆ¶‚ؤ‚ؤŒ¤‹†‚ًگi‚ك‚ؤ‚¢‚éپB“ء‚ةژ÷ڈَچ×–E‚âT, Bچ×–E‚ض‚جgp130ƒVƒOƒiƒ‹‚ھ‚ا‚ج‚و‚¤‚ة–ئ‰u‰“ڑ‚ةٹض—^‚µ‚ؤ‚¢‚é‚ج‚©پH‚ ‚é‚¢‚ح‚±‚جˆظڈي‚ھ‚ا‚ج‚و‚¤‚ب‹@چ\‚إژ©Œب–ئ‰uژ¾ٹ³”ڈا‚ةژٹ‚é‚ج‚©پH‚ئ‚¢‚¤–â‘è‚ً‰ًŒˆ‚·‚ׂŒ¤‹†‚ًگi‚ك‚ؤ‚¢‚éپB‚±‚جƒ‚ƒfƒ‹‚حژ©Œب–ئ‰uژ¾ٹ³‚ًƒTƒCƒgƒJƒCƒ“‚جƒVƒOƒiƒ‹ˆظڈي‚ئ‚¢‚¤ٹد“_‚©‚ç–ئ‰uٹw“IŒ©’n‚©‚ç‚ج‚ف‚ب‚炸پA•ھژqگ¶•¨ٹw“Iژè–@‚ًژg—p‚µ‚ؤ‰ًگح‚·‚邱‚ئ‚ھڈo—ˆ‚é‹Mڈd‚بƒ‚ƒfƒ‹‚إ‚ ‚éپBچ،Œم‰نپX‚جŒ¤‹†ژ؛‚ج’·”N‚ة‚ي‚½‚éƒVƒOƒiƒ‹“`’B‹@چ\‚ج•ھژqگ¶•¨ٹw“IŒ¤‹†‚جŒoŒ±‚ًٹˆ‚©‚µ‚ؤپAŒ¤‹†ژ؛‚ج‘چ—ح‚ًŒ‹ڈW‚µ‚ؤپA‚±‚ج–ئ‰uٹwپAˆمٹw‚ة‚¨‚¯‚éچإڈd—v–½‘è‚إ‚ ‚èپA‚©‚آژ„‚ھ‚Q‚W”N‘O‚ةˆمٹw•”‚ً‘²‹ئ‚µ‚ؤپA–ئ‰uٹw‚ة“¥‚فچ‚ٌ‚إˆب—ˆ‚ج–²‚إ‚ ‚éپAƒZƒ‹ƒtƒgƒŒƒ‰ƒ“ƒX‚ج”j‰َ‹@چ\پA‚·‚ب‚ي‚؟ژ©Œب–ئ‰uژ¾ٹ³‚ج”ڈا‹@چ\‚ً‰ً–¾‚µپAژ©Œب–ئ‰uژ¾ٹ³‚جژ،—أ–@ٹJ”‚ج‚½‚ك‚جٹî‘bڈî•ٌ‚ً“¾‚½‚¢‚ئچl‚¦‚ؤ‚¢‚éپB‚¨‚¨‚¢‚ةگV‹K‚بژل‚¢Œ¤‹†ژز‚جژQ‰ء‚ًٹْ‘ز‚µ‚ؤ‚¢‚éپB
Œ»چفگiچs’†‚ج‹ï‘ج“I‚بŒ¤‹†ƒvƒچpƒWƒFƒN‚ئ‚حˆب‰؛‚ج‚و‚¤‚إ‚ ‚é
‚PپCˆâ“`“I”wŒi‚ئژ©Œب–ئ‰u‚جٹضŒWپA‘£گiˆâ“`ژqپA—}گ§ˆâ“`ژq‚ج“¯’è‚ً–عژw‚·
‚QپCژ©Œب–ئ‰u‚ً‘£گi‚·‚鑼‚ج—vˆِ‚ج‰ًگحپiچ׋غٹ´گُپAHTLV-p40Tax‚ب‚ا)
‚RپC”ڈاپA‹y‚رƒGƒtƒFƒNƒ^پ[‚ةٹض—^‚·‚éچ×–E‚ج“¯’è‚ئپA•ھژqƒŒƒxƒ‹‚ة‚¨‚¯‚éˆظڈي
‚SپC”ڈاˆب‘O‚ة‚¨‚¯‚éˆظڈيپAچ×–E‚جƒŒƒxƒ‹‚©‚çپAƒVƒOƒiƒ‹‚جƒŒƒxƒ‹‚©‚çپiDNAƒ`ƒbƒvپAˆâ“`ژq”Œ»پA“™پj
‚TپCTچ×–E‚جƒlƒKƒeƒBƒuپAƒ|ƒWƒeƒBƒu‘I‘ً‚جˆظڈي
‚UپCƒپƒ‚ƒٹپ[Tچ×–E‚جˆظڈي
‚VپCژ÷ڈَچ×–E‚جˆظڈيپA
‚WپC‹¹‘B‚ة‚¨‚¯‚éTچ×–E•ھ‰»ˆظڈيپAچ×–E‰^“®ˆظڈي
‚XپC”jچœچ×–E‚جˆظڈي
پ@
پ@
پ@
|
|
|
|