|
ニューロトロフィン(NGF, BDNF, NT3, NT4/5)は発生途上においてプログラム細胞死を防止する物質として知られているが、一方で細胞死を誘導することも報告されている。これはニューロトロフィンが2つの異なった種類の受容体を介してシグナルを伝えるためと考えられる。一つはtrk受容体であるが、この受容体の活性化を介して様々な栄養因子様作用が生み出されている。一方、p75受容体もニューロトロフィンをリガンドとする受容体であるが、TNF受容体あるいはCD95(FAS/Apo-1)などの属する受容体ファミリーの一員であり、trk受容体とは極めて異なった性質を持っている。
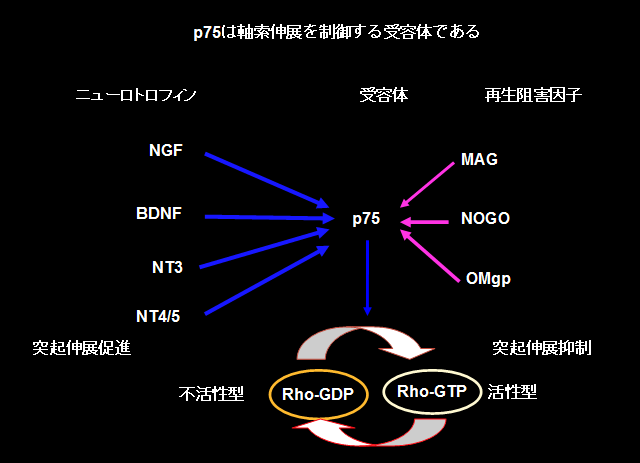
私たちは、p75が発生期の軸索の成長を制御する受容体であることを発見した(Yamashita et al., Neuron, 1999)。ニューロトロフィンがp75に結合すると、small GTPaseであるRhoの不活性化がおこり、それにより突起の伸展が促進されるというメカニズムを明らかにした(図1)。この発見により、p75の機能を明らかにするとともに、Rhoが軸索伸展を制御する重要な細胞内シグナルであるという概念を確立した。
その後、このp75が、ニューロトロフィンのシグナルを伝えるのみならず、myelin associated glycoprotein (MAG)というミエリンの中に存在する糖タンパク質からのシグナルをも神経細胞に伝えていることを見いだした(図1; Yamashita et al., J. Cell Biol., 2002)。MAGは、ニューロトロフィンとは逆に、神経突起の伸展を阻害する作用をもっており、大人の中枢神経の軸索がいったん損傷されると、もはや再生できない原因の一つと考えられている。また突起伸展阻害はRhoの活性化によるものであった。
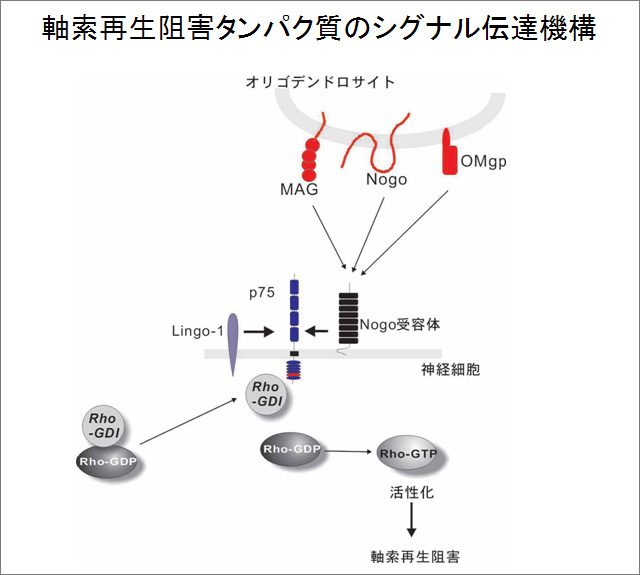
その後、他のグループによってNogoとOMgpという再生阻害タンパク質も、p75を介してRhoを活性化していることが示され、当時見つかっていた中枢のミエリンの中に存在する軸索再生を阻害するタンパク質は全て、p75受容体を介して軸索再生をブロックしていることが明らかになった。私たちはさらに、p75がRhoGDIとRhoとの結合を解離することでRhoを活性化するという詳細な分子機序を明らかにし(図2)、さらにp75の細胞内ドメインに結合し、シグナル伝達をブロックするペプチドを発見した(Yamahsita et al., Nature Neurosci., 2002)。
ニューロトロフィンの刺激により、活性化されたPKAがp75をリン酸化することで、p75が細胞膜のラフトに移動すること (Higuchi et al. EMBO J, 2003)、Gタンパク質およびPKCがP75を介したMAG、Nogoのシグナルを運ぶこと(Hasegawa et al., J Neurosci., 2004)、さらにMAGの下流でRap1が活性化され細胞の生存を制御していること(Taniguchi et al., Cell Death Differ., 2008)などを報告し、p75の多様なシグナル伝達機構とその機能の解明に貢献した。
|
 |
|
|
|
|
|
|
| 参考論文 |
|
- Yamashita, T., Tucker, K. L. and Barde, Y.A. (1999) Neurotrophin binding to the p75 receptor modulates Rho activity and axonal outgrowth. Neuron 24, 585-93.
- Yamashita, T., Higuchi, H., and Tohyama, M. (2002) The p75 receptor transduces the signal from myelin-associated glycoprotein to Rho. J. Cell Biol. 157, 565-570.
- Yamashita, T. and Tohyama, M. (2003) The p75 receptor acts as a displacement factor that releases Rho from Rho GDI. Nature Neurosci. 6, 461-467.
- Higuchi, H., Yamashita, T., Yoshikawa, H. and Tohyama, M. (2003) PKA phosphorylates the p75 receptor and regulates its localization to lipid rafts. EMBO J. 22, 1790-1800.
- Higuchi, H., Yamashita, T., Yoshikawa, H. and Tohyama, M. (2003) Functional inhibition of the p75 receptor using a small interfering RNA. Biochem. Biophys. Res. Commun. 301, 804-809.
- Hasegawa, Y., Fujitani, M., Hata, K., Tohyama, M., Yamagishi, S. and Yamashita, T. (2004) Promotion of axon regeneration by MAG and Nogo through divergent signals downstream of Gi/G. J. Neurosci. 24, 6826-6832.
- Fujitani, M., Kawai, H., Proia, R.L., Kashiwagi, A., Yasuda, H. and Yamashita, T. (2005) Binding of soluble myelin-associated glycoprotein to specific gangliosides induces the association of p75NTR to lipid rafts and signal transduction. J. Neurochem. 94, 15-21.
- Taniguchi, J., Fujitani, M., Endo, M., Kubo, T., Fujitani, M., Miller, F.D., Kaplan, D.R. and Yamashita, T. (2008) Rap1 is involved in the signal transduction of myelin-associated glycoprotein. Cell Death Differ. 15, 408-419.
- Watanabe, T., Ito, T., Inoue, G., Ohtori, S., Kitajo, K., Doya, H., Takahashi, K. and Yamashita, T. (2008) The p75 receptor is associated with inflammatory thermal hypersensitivity. J. Neurosci. Res. 86, 3566-3574.
- Omoto, S., Ueno, M., Mochio, S., Takai, T. and Yamashita, T. (2010) Genetic deletion of paired immunoglobulin-like receptor B does not promote axonal plasticity or functional recovery after traumatic brain injury. J. Neurosci. In press.
|
|
|