細胞生物学
- なぜ腸の上皮細胞の内腔面だけに微絨毛が生えるのか?どうやって神経は軸索を伸ばすのか?
- 腸、神経、免疫細胞の方向性(極性)を決める分子機構には共通点があるのか?
- 動物個体や初代培養系を用いた、生体内に近い状態の細胞の機能解析
- 超解像顕微鏡、電子顕微鏡による微細構造の違いの観察

細胞の向きはどうやってできるのか?という古来からの問いに最新技術で挑む
1)ノックアウトマウス等を用いた、細胞の極性輸送のメカニズムの研究
細胞の極性は細胞の機能に重要な役割を果たしています。上皮細胞は頂端側(apical)、側底側(basolateral)という極性を持つことで、分泌などの機能を果たすことができます。同様に神経細胞も軸索、樹状突起という極性のある構造をとることが神経伝達にとって必須です(図1)。
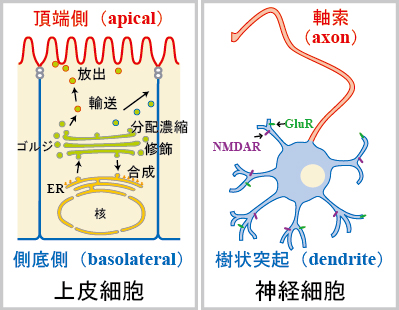 図1
図1
このような極性を持つ細胞においては、色々なタンパク質が、方向性のある輸送(極性輸送)によって目的地に運ばれてその役割を果たしています。この極性輸送は、主に a. ゴルジ体等における輸送小胞へのタンパク質の分配、濃縮 b. 輸送小胞の目的地への輸送 c. 輸送小胞の細胞膜への繋留、融合 で成り立っていると考えられています。
2)細胞の極性輸送のメカニズムを解明するには?
当研究室では、タンパク質の分配や輸送に重要な役割をしていると考えられるタンパク質の遺伝子(SNAREタンパク質、Rabタンパク質等)のノックアウトマウスを作製しています。作製したマウスを様々な細胞生物学的解析法(超解像顕微鏡、電子顕微鏡を用いた形態観察、上皮・神経細胞の初代培養と外来遺伝子の細胞への導入、ライブイメージングを用いた細胞内輸送の解析等)を用い、遺伝子改変マウス個体・細胞において極性輸送の解析を行っています(下記)。 さらに、このような既存分子の解析だけでなく、GST-pulldown法、免疫沈降法などを用いて細胞の極性輸送に重要な新規分子の同定も行っています(下記)。
3)Rab8 ノックアウトマウスの解析で解明されたこと
Rab8は低分子量GTP結合タンパクの1つで、小腸の吸収上皮細胞に多く発現し、basolateral へのタンパク質の輸送に必要と考えられていたタンパク質である。Rab8 を欠損するマウスは吸収上皮細胞で apical に分布する酵素などが細胞内に蓄積して小腸で栄養吸収が殆どできず、生後3~4週後に栄養失調で死亡しました。これらの症状は、微絨毛萎縮症(小腸から栄養が吸収できない病気の1つ)の患者さんの症状と非常に良く似ているため、微絨毛萎縮症の患者さんの小腸で Rab8 の量を調べたところ、Rab8が大幅に減少していることが分かりました。この研究により、腸における栄養吸収のメカニズムの基盤を解明すると共に、腸からの栄養吸収が低下する病気のモデルマウスを作製することが出来ました。 さらに、Rab8がどのような分子機構でapical側への小胞輸送を担うかを解析するため、酵母2ハイブリッド法を用いて、Rab8に結合するEHBP1L1というタンパク質を同定しました。EHBP1L1は膜を曲げて小胞を切り出す際に働くBin1というタンパク質と結合するため、Rab8, EHBP1L1, Bin1が一体となって働いてapical側に向かう小胞を形成すると考えています(図2)。
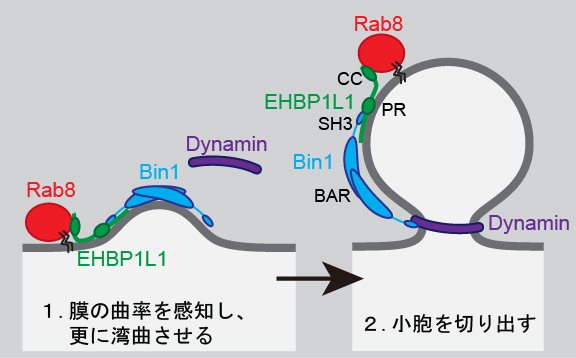 図2
図2
現在、他にapical側への輸送に関与すると思われる遺伝子のノックアウトマウスの作製と解析、及び、EHBP1L1などに結合する分子の探索を行っており、apical側にタンパク質を輸送する分子機構の解明に取り組んでいます。









